|
|
|
RESULTSQualitative Variation
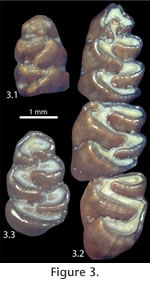
Qualitative variation in the occlusal pattern through ontogeny is the same for the populations from both Inglis 1A and Inglis 1C, with the exceptions specified below. Few individuals in either population exhibit a juvenile pattern, possibly due, at least in part, to the brachydont nature of the teeth that show change in occlusal morphology with relatively little wear and destruction during predation. The symmetrical shape of the anteroconid is the last feature on the m1 to attain the mature wear pattern. After the mature pattern of the m1 is reached, both the associated m2 and m3 develop an enamel pit from the constriction of the second buccal reentrant angle (BRA2; protoflexid of
Peláez-Compomanes and Martin 2005) anterolabially by the anterior cingulum (Figure 3.2). The anterior cingulum is low on the tooth and therefore does not close off the BRA2 in ontogenetically-younger individuals. Among the Inglis 1C specimens, each instance of an m3 with a BRA2 pit occurs with an m2 having the same feature.
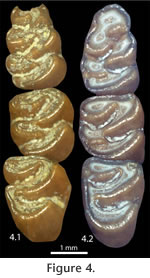
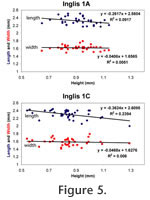
With progressive wear all BRA2 enamel pits on the m2 and m3 are worn away, and the lingual openings in all lower molars become tightly constricted (Figure 3.2, Figure 4.1). This constriction is least developed in the second lingual reentrant angle (LRA) of the m1 (because this feature is deeper than other lingual reentrant angles) and the buccal reentrant angles of the m1 (because of asymmetrical wear of the teeth). In the most advanced wear stage observed in S. curtisi, the lingual reentrant angles are closed, or nearly so (Figure 4.2); these individuals still retain the buccal reentrant angles on the m1 and the first buccal reentrant angle on the m2 and m3. The connection between the anteroconid and metaconid is along the labial edge of the tooth. Enamel pits from the constriction of the BRA2 never developed on the m1. Quantitative VariationThe age cohorts of individuals of S. curtisi from Inglis 1A and Inglis 1C correlate closely with the height of the m1 (as measured from the linea sinuosa to the metaconid; Table 1), which is reduced as a result of increased wear in ontogenetically older individuals. Two exceptions are specimens from Inglis 1A: UF 22024 does not yet have the BRA2s constricted into enamel pits (cohort III in Table 1) and has a shorter than expected height of 0.79 mm; and UF 22044 has a taller than expected height of 0.90 mm for an individual with the BRA2 enamel pits already eroded away (cohort V in Table 1).
The number of roots on the m1 was shown previously to vary within samples of fossil cotton rats, and this variation has been used, in part, for species and species-group identifications (Martin 1979). Previous examination of S. curtisi from Inglis 1A documented three roots in 14 of 16 m1s, with the remaining two m1s having four roots (Martin 1979). My study includes specimens not then known to Martin, plus the larger sample from Inglis 1C. Table 2 shows a higher occurrence of four-rooted m1s in S. curtisi than previously recorded; more notable is the rare occurrence (not previously noted in published literature) of specimens with only two roots in both the Inglis 1A and Inglis 1C samples. I compared the abundance of roots to measurements of the m1. Within each of the Inglis 1A and Inglis 1C samples, there was no significant difference in the length, width, or height when specimens were pooled according to their number of roots (Table 2). |
|