|
|
|
MORPHOLOGY AND RELATIONSHIPS OF SLAUGHTERIA ERUPTENS
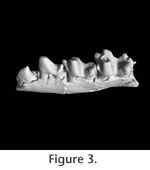
One reinterpreted salient character of S. eruptens is the lack of molarization of the posterior premolars, in contrast to eutherian mammals (Kielan-Jaworowska et al. 2004).
Slaughter (1968b) referred what he identified as submolariform premolars from the Trinity Group to Eutheria and later, by inference, to Pappotherium, which he considered a eutherian (Slaughter 1981). A submolariform lower premolar figured by
Slaughter (1968b) differs in morphology from the dp4 S. eruptens. It either derives
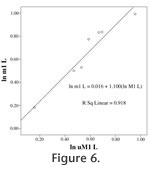
Molar length has been used as a character in phylogenetic reconstructions of Cretaceous eutherian mammals (Archibald and Averianov 2006). The length of the newly interpreted lower m1 of S. eruptens closely matches first molars of Eomaia scansoria (Ji et al. 2002) and Prokennalestes minor (Kielan-Jaworowska and Dashzeveg 1989). Proposed Taxonomic AssociationsPairing of isolated upper and lower teeth of Trinity Group tribosphenidan mammals has been attempted since Patterson (1956) described, but did not name the first sample of these teeth. Despite new collections, little consensus has resulted (Slaughter 1965, 1971; Turnbull 1971; Butler 1978). Upper teeth of Trinity Group tribosphenidans are differentiated primarily on morphology of the stylar shelf and protocone. The first tribosphenidan taxa named from the Trinity Group (Pappotherium pattersoni Slaughter 1965 and Holoclemensia texana Slaughter 1968a) were based on upper molar teeth. These two mammals differ notably in size as well as stylar cusp development. Other Trinity Group tribosphenidan taxa that were named, based upon upper teeth, include Comanchea hilli (Jacobs et al. 1989), Atokatheridium boreni (Kielan-Jaworowska and Cifelli 2001), and Oklatheridium szalayi (Davis et al. 2008). The four remaining taxa are named from lower teeth or jaws. Patterson (1956) recognized three distinct morphologies of therian lower molars (structural types 1-3) (Table 1). Based upon additional material, Slaughter (1965) added three more morphotypes (4-6) to Patterson's previous three. Several of these morphotypes were described originally from single teeth. In his first paper on the subject, Slaughter (1965) noted that his type 6 lower molar (one molar in a jaw fragment; SMU 61728) was of the size expected to match the uppers of P. pattersoni. This specimen was subsequently designated the type of a new genus and species Trinititherium slaughteri (Butler 1978). Slaughter (1971) described a left lower mammalian jaw with four teeth (SMU 61992), and tentatively allocated it to P. pattersoni based upon size and probable occlusal relationships. He did not comment on its relationship to the morphotypes of Patterson (1956) or Slaughter (1965). Turnbull (1971) assigned all the teeth of Patterson's types 1 and 2 and Slaughter's type 5 to Holoclemensia. One small tooth (PM 922) formed the basis for Patterson's (1956) morphotype 3. This tooth, along with Slaughter's types 4 and 6 (6 would become the type of T. slaughteri), and some more recent discoveries, were referred by Turnbull (1971) to Pappotherium. In contrast, Butler (1978) associated Patterson's type 2 lower molars with Pappotherium (and types 1, 4, 5 with Holoclemensia), but he argued that molariform teeth in the jaw SMU 61992 differed from other type 2 molars and named the lower jaw Slaughteria eruptens. Butler (1978) also believed that type 3 teeth (including PM 922) resembled Kermackia texana (Slaughter 1971), a taxon described from lower teeth nearly simultaneously with Turnbull's (1971) paper. Slaughteria eruptens was regarded by Butler (1978) as smaller than teeth that he referred to Pappotherium. Thus, P. pattersoni, which is based upon upper molars, has been variously associated with lower molars of morphotypes 2, 3, 4, or 6 (two of which are now designated as separate taxa), and S. eruptens has been regarded as associated lowers of P. pattersoni or recognized as a distinct taxon (Table 1). The only area of real agreement in previous studies is that types 1 and 5 teeth are Holoclemensia (Turnbull 1971; Butler 1978). Tooth size alone clearly differentiates at least two groups of lower molars from the Trinity Group (Figure 5, Table 1; Jacobs et al. 1989; Kobayashi et al. 2002). Tooth Size CorrespondenceOur objective is to constrain quantitatively the size of the lower molars that are compatible with the holotype and unequivocally referred upper molars of P. pattersoni. We also test the associations that have been made of isolated lower teeth with the holotype and unequivocal upper molars of H. texana. Within mammalian species, it is well established that cheek teeth exhibit patterns of size correlation (correlation fields of Kurten 1953). Adjacent and occluding pairs of teeth typically exhibit the strongest correlations, especially within fields such as premolars or molars (Kurten 1953; Van Valen 1962; Gould and Garwood 1969, Gingerich and Winkler 1979; Szuma 2000). A model for estimating opposing tooth sizes in Early Cretaceous mammals is limited by the lack of population size samples of skulls with associated dentitions. However, given the limited range of tooth morphologies found in Cretaceous mammals with tribosphenic molars and their similar dental formulae, it is possible to predict that corresponding pairs of occluding upper and lower molars (especially M/m1 M/m2) across a range of species will also be constrained to strong size correlation in order to maintain occlusal relationships. Assuming robust length correlations between occluding molar teeth, an estimate of lower molar length based upon a given upper molar length can be made using regression analysis. Archibald and Averianov (2006) use differences in upper M1/lower m1 length ratios as phylogenetic character states in analyzing Asian Cretaceous mammals. Such varying proportions could confound the assumptions in the following regression analysis. However, systematic differences in upper/lower molar length ratios would be apparent in the following analysis in scatter plots and measures of length relationships (see below).
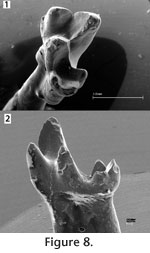
There is clearly a group of lower molars from the Trinity Group (Figure 5) that are smaller than the m1 of S. eruptens and other teeth of similar morphotype (e.g., PM 948;
Figure 9). Small lower molars that are referred to
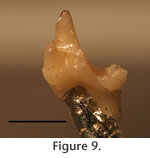
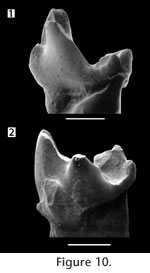
Deciduous Teeth. Recognition of a deciduous tooth in S. eruptens affords an insight not previously available; the diagnosable morphology of a molariform deciduous premolar in Trinity Group tribosphenidan mammals. Important characters, in addition to small size and narrowness, include the relatively low, anteroposteriorly elongate trigonid with a small paraconid rotated to the midline of the tooth. These characters were noted in other deciduous teeth of Cretaceous mammals (Butler 1977). Several teeth from the Trinity Group fit this description including PM 922 (Figure 10; Patterson's 1956, "type 3") and Kermackia texana, and invite further examination of the status of these specimens to identify them as permanent or replacement teeth. More than one taxon of smaller therians exist within the Trinity Group, but how many remains open to question. It is also not clear whether any "molarized" premolars from the Trinity Group (Slaughter, 1968b) represent permanent teeth. |
|