|
|
|
RODENT PALEONTOLOGYThe following is a list of rodent taxa identified from Golliher B:
Family Sciuridae
Family Geomyidae
Family Arvicolidae
Family Cricetidae
Family Dipodidae The Golliher B rodent assemblage is compared with a few radiocarbon-dated rodent assemblages from southern Nebraska, Kansas and Oklahoma in Table 1. Dates for these localities are provided in Table 2. The rodents of an earlier, cold-adapted fauna (Cudahy; Paulson, 1961), those of the Jinglebob l.f. (Hibbard 1955) and the modern rodent fauna of Meade County are also listed. The Jinglebob l.f. of Meade County, Kansas, lacks a radiometric date, and its fauna is not sufficient to establish a precise age, though it was most recently allocated to the "Wisonsinan" by Miller (1975). The combined presence of Microtus pennsylvanicus, M. ochrogaster and the extinct Synaptomys australis certainly suggest a late Pleistocene (Rancholabrean) age, and likely younger than assemblages such as Butler Spring and Mt. Scott (Schultz 1969). The extralimital record of Oryzomys palustris also suggests that Jinglebob was deposited during a relatively warm period, and it provides a useful contrast both for the earlier Cudahy l.f. with its host of extinct arvicolids and the radiocarbon-dated sites in Table 1 with their boreal affinities. Those species of climatic importance from Golliher B are considered individually below.
Family Geomyidae Material examined. FHSM 17027, Lp4, LM3; FHSM 17028, Lp4, L upper molar. Discussion. Thomomys species are easily diagnosed by upper incisors without grooves and p4 with V-shaped reentrant folds lacking cement. The lower molars also tend to be asymmetrical, or "tear-dropped" shaped, and have enamel on both leading and trailing edges. Thomomys first appears in the Cudahy l.f. at 0.64 Ma (Paulson 1961), associated with other species indicating a cold temperate climate, and it is present in all later Pleistocene faunas subsequent to Jinglebob (Hibbard 1950). The Jinglebob l.f. was first considered by Hibbard (1950) to be of "Sangamon" age, but it was later placed by Miller (1975) in a later, colder period of the late Pleistocene. Without radiometric dates the age of this site cannot be secure, and despite Miller's (1975) analysis, the presence of the rice rat Oryzomys palustris and southern bog lemming, Synaptomys australis, suggest that if the fauna accumulated during the latest Pleistocene, it was probably during a warm interstadial period. Schultz (1969) tentatively identified the northern pocket gopher, Thomomys talpoides, from the late Pleistocene Robert l.f., and based on zoogeographic considerations and other disjunct species in the various Meade Basin assemblages indicating cooler conditions, this is the most likely species to have been present in Kansas during the late Pleistocene. The northern pocket gopher today has outlier populations in eastern Nebraska and eastern Colorado. Nevertheless, at some point it would be helpful if dental or mandibular characters could be identified to distinguish between T. talpoides and its southern relative T. umbrinus.
Family Arvicolidae
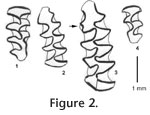
Discussion. Like Thomomys, the meadow vole is a ubiquitous presence in Meade Basin assemblages of late Pleistocene age. M. pennsylvanicus can be identified given adequate sample sizes of m1, M2 and M3. An M2 with incipient or closed triangle 4 (T4) is particularly diagnostic (Zakrzewski 1985; Davis 1987; Semken and Wallace 2002), although this feature does appear occasionally in other species (Bell and Repenning 1999). Ten of the 14 M2s from Golliher B examined in the size range of M. pennsylvanicus possessed this extra triangle. In a large sample of Microtus M2s from the upper Ambystoma faunule of the Jones l.f. of Meade County, Kansas (26,700 + 1500 and 29,000 + 1300 years B.P.), Davis (1987) also noted some specimens without T4, and he speculated on the presence of another Microtus other than M. pennsylvanicus and M. ochrogaster, both of which he had identified from the Ambystoma faunule. Stewart (1987) reported Microtus montanus from a few late Pleistocene sites in northern Kansas and Nebraska. Conceivably, another Microtus species could be present in the Golliher B l.f. in addition to M. pennsylvanicus, but it is also possible that there is either 1) an evolutionary tendency towards greater expression of T4 through time in the meadow vole or 2) geographic polymorphism within extant populations that has not been quantified, or both. Davis (1987), building on preliminary data presented by Semken (1966), documented clinal variation in the number of triangles on m1 in extant North American M. pennsylvanicus, with the most complex m1s found on specimens from the Central Great Plains. This complexity evolved through the late Pleistocene. Perhaps polymorphism and evolutionary change can also be documented for the M2. Measurements of a sample of M. pennsylvanicus m1s from Golliher B are presented in Table 3. Davis (1987) documented a shift in meadow vole m1 morphotype diversity on the Central Great Plains, from 100% 5T forms in the late Pleistocene Sandahl l.f. through 30.1% 5T forms in extant populations. Dates are not available for the Sandahl and Mt. Scott l.f.s, but they are probably older than the Jones l.f. and definitely younger than the Cudahy l.f. (Paulson 1961) at 0.64 Ma. Part of Davis's (1987, table 8) summary is reproduced chronologically in Table 4, with comparable data for a sample from Golliher B (some of Davis's totals do not add to 100% because he also counted 7T forms separately and additionally has an undefined category labeled "other"). These data show that the switch from 0.67/0.33 (5T/6T) to 0.33/0.67 (5T/6T) populations occurred in Kansas in a remarkably brief 1500 year period between 12,500-11,000 radiocarbon years ago. The modern proportional representation is seen in the Robert l.f. at 11,000+ 390 radiocarbon years B.P., just at the beginning of the Younger Dryas (YD) event. It is tempting to speculate that the dramatic YD climatic environment played a part in selecting for the final m1 configuration in M. pennsylvanicus on the Central Great Plains, but if so the YD event did not similarly influence meadow vole populations elsewhere within the species' geographic range. Davis's (1987) figure 12 shows that closure of T6 on m1 in extant M. pennsylvanicus is most common on the Central Great Plains; populations to the north, east and west more commonly display five closed triangles on m1. Martin and Prince (1989) showed that the proportion of 5T m1s in M. pennsylvanicus from Bell Cave in northern Alabama averaged 79% between about 26,500-11,800 radiocarbon years B.P., and Gordon (1999) reported an average of 93% 5T forms in 13 levels from Yarbough Cave, Georgia, ranging from about 23,880-16,500 radiocarbon years B.P. Although the meadow vole resides today farther north in the southern Appalachians than either Bell Cave or Yarbrough Cave, from Davis's (1987) and Gordon's (1999) data, we could expect the modern eastern proportion to range between 70-100% 5T forms. Thus, it appears that eastern meadow vole populations retained the ancestral proportion of 5T morphotypes through the later Pleistocene into modern time, as Semken (1966) had earlier hypothesized. Davis (1987) speculated that the higher complexity of Great Plains meadow vole m1s was correlated both with greater tooth length and foraging on a higher proportion of harsh grasses. Martin and Prince (1989) found no correlation between m1 length and closure of T6 in the Bell Cave and Yarbrough Cave samples of M. pennsylvanicus, and as there has been no quantitative assessment of diet and dental size or complexity in meadow voles, we cannot be certain of the factors contributing either to variation in the modern populations or the rapid shift from 5T to 6T morphotypes on the Great Plains during the latest Pleistocene. The earliest appearance of M. pennsylvanicus in the Meade Basin is in the late Pleistocene (Rancholabrean) Adams l.f. in Meade County (Schultz 1969), and it is present in the region until it becomes locally extinct during the Holocene. In the Central Great Plains and nearby regions, the meadow vole is found today only in extreme northern Kansas and at higher elevations in Colorado and northern New Mexico, about 300 km from Meade County. Its local extirpation in southwestern Kansas appears to be roughly synchronous with the Holocene immigration of the hispid cotton rat, Sigmodon hispidus, both events roughly correlated with the beginning of the current interstadial warm period. Among many first lower molars of M. pennsylvanicus recovered from Golliher B, there were three demonstrating an unusual morphology. In these specimens, a prism fold filled with cement is present (Figure 2). This feature is characteristic of Mimomys species with rooted molars. Rather than representing an ancient population preserving archaic characters, we suspect these specimens represent individuals in which part of the ancestral epigenetic system became functional once again. Zakrzewski (1985) also reported a prism fold in an m1 of extant M. longicaudus.
Microtus (Pedomys) parmaleei, new species Holotype. FHSM 17023, Lm1. Paratypes. FHSM 17024, part Lm1; FHSM 17025, part Lm1; FHSM 17026, 2 Rm2, 1L and 1R M2; 2R and 1L M3. Referred specimens. The fossil material listed as Microtus ochrogaster in Martin (1991) from the following localities in the eastern United States: Yarbrough Cave, Georgia (BC 291 -- 4 Lm1, 1 R mandible; BC 292 – 2 L mandibles, 6L and 2 R m1s; BC 293 – 1 R mandible). Cheek Bend Cave, Tennessee (UT 135-79 – 9 m1s; UT 113-79 – 5 m1). Clark's Cave, Virginia (CM 24524 – 1 L and 1 R mandible, 6 L and 3 R m1s). Natural Chimneys, Virginia (CM 7505 – 3 R and 5 L mandibles; 18 R and 13 L m1s). The Microtus ochrogaster material listed by Schultz (1969) from the Robert l.f. (UM-K1-57) of Meade County, Kansas: UMMP 46016, 41469 (two right mandibles), UMMP 41470 (isolated molars). Locality and horizon. Level B of the Golliher sequence along Sand Creek, Meade County, Kansas, 12,510 + 80 14C years B.P. (latest Pleistocene). Diagnosis. A small Pedomys characterized by a combination of m1s with primitive and advanced m1 morphotypes; BRA4 and LRA5 are often absent to poorly developed and devoid of cementum. Separable from modern M. ochrogaster by smaller size and greater proportion of primitive m1 morphotypes. Differs from M. ochrogaster /llanensis on basis of considerably smaller size. Discussion. Among the numerous remains of arvicolid rodents from Golliher B, the vast majority of which were identified as Microtus pennsylvanicus, the meadow vole, were 10 molars from a diminutive member of the subgenus Pedomys. This small vole presents an enigma, as two of three m1s demonstrate a relatively primitive morphology compared to that of the modern Microtus (Pedomys) ochrogaster from Meade Kansas as well as some prairie vole specimens recovered from older Wisconsinan sediments in Meade County (e.g., Butler Spring, Schultz 1969). In size and morphology the small Pedomys from Golliher B approximates exactly a series of Pedomys samples from Wisconsinan localities in the eastern United States that Martin (1991) identified informally as Parmalee's steppe vole, described in the literature as a chronomorph of M. ochrogaster, M. o. /parmaleei (Martin et al. 2008). The size of the single complete m1, FHSM 17023 (Figure 2), is 2.31 mm long by 0.88 mm wide. In morphology, two of the three m1s (FHSM 17023, 17024) display three closed triangles, T4-5 confluent and a simple anteroconid (acd) in which buccal reentrant 4 (BRA4) is undeveloped and lingual reentrant 5 (LRA5) is shallow and lacks cement. Both reentrants are developed in FHSM 17025; BRA4 is filled with cement but LRA5 is not. This sort of variation is common in the M. o. /parmaleei from such localities as Yarbrough Cave, Georgia, Clark's Cave, Virginia, Natural Chimneys, Virginia, and Cheek Bend Cave, Tennessee (Martin 1991). The specimens reported by Schultz (1969) from the Robert l.f. as M. ochrogaster are small and are also referred to Parmalee's steppe vole. Van der Meulen (1978) proposed that the extant prairie vole, Microtus ochrogaster, evolved from the early Pleistocene M. guildayi, through an intermediate species M. llanensis that Hibbard had named from the Cudahy l.f. (Hibbard 1944). To reflect a more biological nomenclature of phyletic series, Martin (1995) synonomized M. guildayi with M. pliocaenicus and M. llanensis with M. ochrogaster, reducing the taxa to the informal chronomorphs arranged in the following lineage: M. p. /guildayi – M. o. /llanensis – M. o. /ochrogaster. He also presented a preliminary phylogenetic analysis for Pedomys. While correct in essence (inasmuch as a binomial system can ever represent a complex evolutionary population dynamic), the evolution of modern prairie vole populations was probably not that simple. The prairie vole today exhibits a reverse Bergmann's response, with the smallest populations residing in southern Canada and the northern United States in primarily boreal habitat. The diminutive northern populations are recognized as the subspecies M. o. minor. As noted above, samples of small prairie voles are commonly encountered in late Wisconsinan Appalachian fossil assemblages noted for their boreal rodent indicators, including Microtus xanthognathus, Phenacomys intermedius and Synaptomys borealis (Guilday 1962; Guilday et al. 1977; Martin 1991). Occasionally, samples of larger individuals are recorded from Appalachian faunas at the same time (Martin 1991). First lower molars of the small individuals include specimens that are basically indistinguishable from those of M. llanensis, which are themselves only slightly advanced over M. p. /guildayi. Nevertheless, also among both the small Wisconsinan Pedomys and M. llanensis are some specimens indistinguishable from modern M. ochrogaster (Martin 1991, 1995). To make matters more interesting, as Martin (1991) demonstrated, the small extant northern M. o. minor have the most complex m1s among living M. ochrogaster. So how do we interpret these Wisconsinan populations of small size and relatively primitive morphology? There are at least three possibilities: 1) the small animals represent a distinct, dwarf, sibling species (M. parmaleei) of M. ochrogaster with cold steppe and boreal affinities that became extinct throughout its range prior to the Holocene, 2) the small animals represent dwarf M. o. /llanensis that gave rise to M. o. minor, and as climate changed at the end of the Pleistocene they retreated to northern refugia and finalized the push for complex dentitions, or 3) the extant Microtus ochrogaster minor is actually a distinct species, M. minor, descended from dwarf M. o. /llanensis (= M. o. /parmaleei). In scenarios 1 and 3, it is conceivable that van der Meulen's M. p. /guildayi was ancestral to the diminutive species (M. parmaleei or M. minor), and that M. ochrogaster evolved from larger-sized Great Plains M. pliocaenicus through M. o. /llanensis. First lower molars of M. p. /guildayi are decidedly smaller than those of M. llanensis, thus at least giving some credence to this idea. Currently, it is difficult to test these hypotheses. A modern genetic analysis comparing M. o. minor and other M. ochrogaster subspecies would be helpful. Our current evaluation is that the samples of small Pedomys m1s with a high percentage of archaic morphotypes that appear in late Wisconsinan assemblages, known by independent criteria to have existed during much colder climatic periods relative to modern time, represent a distinct species M. (Pedomys) parmaleei. In the Meade Basin it replaced earlier, large-sized Pedomys referable to M. ochrogaster, and was later replaced by larger-sized M. ochrogaster in the modern rodent community of the same region. The same interpretation can be applied to late Pleistocene Appalachian assemblages. Consequently, M. parmaleei can be added to a small list of rodent species that generally indicate colder environments. In the east, this coterie would likely be representative of boreal or sub-boreal conditions, whereas on the Central Great Plains the environment probably ranged from boreal forest in the north to cold steppe in the south (southern Kansas and northern Oklahoma and Texas).
Family Dipodidae Material examined. VP 17029, 1L and 1R m1. Discussion. Jumping mice have been present in the Meade Basin since the earliest Pliocene (Fox Canyon; Hibbard 1950; Klingener 1963). Because their populations are not high in extant rodent communities, they are rarely common in fossil deposits, and their absence from any given Meade Basin assemblage is more likely an artifact of sampling rather than a climatic signal. Klingener (1963), Martin (1989, 1994) and Ruez and Bell (2004) reviewed the record of North American Pliocene and Pleistocene zapodids and presented characters used to distinguish the modern and extinct species. Characters of the first lower molar serve to distinguish the Zapus princeps (= Z. trinotatus) and Z. hudsonius groups. In the former, the anteroconid is a simple cone composed of a dentine island surrounded by an external enamel border. In the latter group, including the extinct Z. rinkeri, Z. sandersi (= Z. burti) and Z. syksae, as well as in extant Z. hudsonius, there is either a deep anteromedian fold (most species) or a central, isolated enamel atoll (Z. syksae). The two m1s present clearly demonstrate the Z. princeps morphology, and the Golliher B Zapus is referred to that species. Zapus princeps has also been identified from three late Pleistocene sites in northern Kansas and southern Nebraska (Stewart 1987). Schultz (1969) reported Zapus hudsonius from the latest Pleistocene Robert l.f., but the only specimen recovered was a mandible with m2, and as the Robert l.f. has other mammalian species indicating a cooler climate (Schultz 1969; pp. 71-72), it may be that this specimen represents Z. princeps. Z. princeps has a modern distribution very similar to that of Thomomys talpoides, Microtus pennsylvanicus and Microtus ochrogaster minor; it is confined to boreal habitats at northern latitudes or at higher elevations in the Rocky Mountain chain to the south, in central Colorado and northern New Mexico. |
|