|
|
|
SYSTEMATIC PALEONTOLOGY
Family HERPETOTHERIIDAE Trouessart, 1879
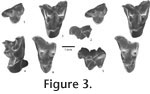
Referred specimens. From NMMNH locality L-6398, NMMNH P-53926, partial left m?; 57342, partial left m4. From NMMNH locality L-7583, 59286, left dP3; 59293, right m2 or 3; 59303, left M2; 59304, left M3; 59345, left M4; 59378, right M1; 59389, left m2 or 3; and 59503, left M4. Horizon and locality. NMMNH localities L-6898 and 7583, Pantolambda cavirictum – Mixodectes pungens Zone (Williamson 1996), Middle Torrejonian (To2) in age (Lofgren et al. 2004; Figure 2). Etymology. Named after Zerina Johanson for her contributions to Cretaceous and Paleogene metatherian studies. Diagnosis. Over 10 percent larger than Swaindelphys cifellii in most dimensions and over 15 percent smaller than S. encinensis sp.nov. in most dimensions. Description. All specimens referred to Swaindelphys johansoni consist of isolated teeth. A single partial tooth is tentatively identified as a dP3 of S. johansoni (Figure 3.1, 3.3). It is missing the distobuccal corner of the tooth. It is smaller than permanent upper molars referred to S. johansoni, but is similar in size relative to the permanent upper molars of other Paleogene metatherians such as Peradectes louisi (Crochet 1979) and Amphiperatherium giselense (Heller 1936) (Crochet 1980, figures 6 and 43, respectively). However, the dP3 of Amphiperatherium goethei (Crochet 1979) and A. maximum (Crochet 1979) are markedly larger relative to upper molars (Crochet 1980, figures 88 and 105, respectively). The protoconal lobe is rounded lingually with the conules placed symmetrically near the protocone on either side. The paraconule is larger than the metaconule. The paracone and metacone are subequal in height. The metacone is wider. The centrocrista is deflected buccally, with a shape resembling an obtuse inverted V. The parastylar lobe is relatively larger than those described for other Cretaceous and Paleogene metatherians. It projects mesially and is mesiolingually expanded so that the mesial margin of the tooth is deeply embayed between the parastylar and protoconal lobes. The intersection of these two lobes describes a nearly right angle at the mesial margin of the tooth. Stylar cusp A is positioned at the mesial extremity of the parastylar lobe and is lingual to a line drawn between the paracone and metacone. A smaller stylar cusp, stylar cusp B (stylocone) is mesial to the paracone, positioned on a line drawn through the paracone and metacone. A low preparacrista extends from the mesial base of the paracone to stylar cusp B. The stylar shelf is narrow buccal to the paracone and expands buccal to the metacone. It supports several stylar cusps. A cusp buccal to the intersection of the postpara- and premetacristae is interpreted to represent stylar cusp C. It is larger than stylar cusp B and smaller than stylar cusp A. It is nearly conular, but mesiodistally elongate. A diminutive cusp is positioned close to and distal to stylar cusp C. Discussion. Nearly all specimens referred to Swaindelphys johansoni were collected from a single locality, NMMNH locality L-7583. Two additional specimens, both fragments of lower molars were collected from L-6398 and are tentatively referred to this taxon. Both of these localities are within the Pantolambda cavirictum – Mixodectes pungens Zone (Williamson 1996) and are Middle Torrejonian (To2) in age (Lofgren et al. 2004). — The upper dentition is represented by several isolated teeth. All four upper molars are significantly larger than the respective teeth of Swaindelphys cifellii (Johanson 1996a, table 1), and there is no overlap in tooth dimensions (Table 2). A dP3 referred to S. johansoni is one of the few that has been described for any Cretaceous or Paleogene metatherian taxa. A dP3 has been described and illustrated for several of these, including Alphadon rhaister (Clemens 1966, figure 11), Didelphodon vorax (Clemens 1966, figure 51), Protolambda hatcheri (Clemens 1966, figure 28), Leptalestes krejcii (Lillegraven 1969, figure 23.5), Glasbius twitchelli (Archibald 1982, figure 49a-b), "Peradectes" cf. "P." pusillus (Archibald 1982, figure 44a-b), Peradectes californicus (Rothecker and Storer 1996, figure 1O), Herpetotherium sp. cf. H. marsupium (Rothecker and Storer 1996, figure 1A), and numerous Paleogene European metatherians (see Crochet 1980). An additional dP3 of a Cretaceous metatherian was described and illustrated by Eaton (1999, figure 3E) as "Pediomys sp." based on an isolated tooth, UMNH VP 6894 (listed as UMNH VP 6594 by Eaton et al. (1999), from the Campanian Wahweap? Formation of Utah. Later, Eaton (2006, figure 13A) described this tooth as an M1 of "Pediomyid gen. and sp. indet." We agree with Davis (2007), that this tooth is a dP3 of "Pediomys sp." A number of the taxonomic identifications of other taxa reported above were considered doubtful or tentative. Regardless, these teeth share a similar overall shape in which the tooth is longer than wide, the protocone is relatively small with closely spaced conules, the parastylar lobe projects mesially, the paracone and metacone are widely spaced, and the stylar shelf is reduced or missing mesially and buccal to the paracone. Among these taxa, salient differences are the relative development of the stylar shelf buccal to the paracone, relative size and development of the parastylar lobe, the relative size and placement of stylar cusp B, which is absent from several taxa, and the relative size and placement of stylar cusps C and D. The stylar shelf of the dP3 is reduced relative to the metastylar lobe in all metatherian taxa. It ranges in development from being present but narrow (e.g., Turgidodon rhaister and Glasbius twitchelli), present only as an ectocingulum on the buccal face of the paracone (e.g., Peradectes californicus), to being completely absent (e.g., Swaindelphys johansoni). The tooth here referred to Swaindelphys johansoni most closely resembles a fragmentary dP3 described by Archibald (1982) and tentatively referred to Peradectes" cf. "P." pusillus. As in S. johansoni, it has a relatively greatly expanded parastylar lobe. Moreover, the parastylar lobe is expanded mesiolingually so that stylar cusp A which occupies a position near the mesial apex of this lobe, is positioned lingual to a line drawn through the paracone and metacone. A similar condition is observed for a tooth, UALP 2872 that Lillegraven (1969) doubtfully referred to Leptalestes krejcii. However, in both taxa, the parastylar lobe is not as expanded as in S. johansoni and both also lack a stylar cusp B. Clemens (1966), in describing the dP3 of Turgidodon rhaister and Archibald (1982) in describing a dP3 that he tentatively referred to G. twitchelli, identified the stylar cusp immediately distobuccal to the paracone as the stylar cusp B and reported that stylar cusp C was absent in these teeth. It is much more likely that the stylar cusp B is absent in these specimens and the cusp immediately mesiodistal to the paracone is stylar cusp C. As in Swaindelphys johansoni, a diminutive stylar cusp B is present mesiobuccal to the paracone in the dP3 of P. californicus.
Swaindelphys encinensis sp. nov.
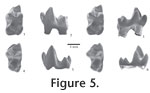
Peradectes? n. sp. B. Williamson and Lucas, 1993 p. 120; Williamson, 1993 p. 101, 1996 p. 34. Holotype. NMMNH P-21594, right partial maxilla with M3 from NMMNH locality L-2685. Referred specimens. From NMMNH locality L-312, NMMNH P-00060, right m1; 01972, left m2 or 3; 57344, left M2; 58843, left partial M2; 58888, left m2 or 3; 58921, left m2 or 3; 58924, right m2 or 3; 58930, partial left m2 or 3 (talonid); 58948, left m2 or 3; 58970, right m1; 58982, left m4; 59004, left partial M4; 59023, right M1; 59028, partial left M3; and 59256, partial left M4. From NMMNH locality L-1287, NMMNH P-54129, left M4. From NMMNH locality L-1486, NMMNH P-18587, right M3. From NMMNH locality L-6261, NMMNH P-53927, partial left m?; 53932, partial right lower molar; 53933, partial right lower molar (partial trigonid); 53935, partial right lower molar (talonid). From NMMNH locality L-6249, NMMNH P-48543, right partial M1 or 2. From NMMNH locality L-6282; NMMNH P-52885, partial right dentary with m1-2. From NMMNH locality L-6315, NMMNH P-53928, right m1. From NMMNH locality L-6898, NMMNH P-57342, partial left m4. Horizon and locality. Numerous localities of the Mixodectes pungens zone (Williamson 1996), all located at the East and West Flanks of Torreon Wash, Nacimiento Formation, San Juan Basin, New Mexico, late Torrejonian (To3) in age (Lofgren et al. 2004). Etymology. Named for Ojo Encino, the area from which the type and referred specimens were collected. Diagnosis. Larger than Swaindelphys johansoni, over 15 percent larger in most dimensions; upper and lower molar cusps more inflated and upper molar protocone more mesiodistally expanded; M4 differs from S. johansoni in possessing a less buccally expanded metastylar lobe and in lacking an ectoflexus. Description. As in other Paleogene marsupials, M1 of Swaindelphys encinensis is relatively less transverse than M2 or 3 and the stylar shelf is reduced mesially, giving it a canted appearance in occlusal view. Also, the protocone is less compressed mesiodistally. The M1 has only a shallow ectoflexus (Figure 4.1). It is progressively deeper in M2 (Figure 4.2) and M3 (Figure 4.3-4.4). In M3 the ectoflexus is relatively deep and smoothly indented, centered on stylar cusp C. In all upper molars the paracone is smaller than metacone. The para- and metacones have nearly flat buccal walls. The metacone has a slight expansion at its base buccally, giving this area a modest convexity. The paracone and metacone are separated by a deep and narrow cleft. The preparascrista intersects with stylar cusp B (= stylocone) in M1 and 2, but merges into the stylar shelf lingual to a position that is medial to the apex of stylar cusp B in M3. There is no contribution to the preparacrista from stylar cusp B in M3. Stylar cusp B is the largest of the stylar cusps, followed by stylar cusp D, stylar cusp C, and finally, stylar cusp A. Stylar cusp A is circular and subequal in height to stylar cusp B. A crista extends mesially from stylar cusp B, but a distinct notch separates this crista from stylar cusp A. Stylar cusp C is situated at the deepest part of the ectoflexus on the ectocingulum so that the ectoflexus is centered on stylar cusp C. Cusp D is represented by a single elongate and oval shape in NMMNH P-18517, but appears to be represented by several smaller cusps along the margin of the stylar shelf in NMMNH P-21594 (Figure 4.3-4.4). In unworn teeth, the paraconule is smaller than the metaconule. The postmetaconule crista terminates below the distolingual base of the metacone. The preparaconule crista extends buccally to the lingual base of stylar cusp A. A postparaconule crista is indistinct or absent. A premetaconule crista is present and extends bucally from the metaconule to the base of the metacone. The M4 (Figure 4.5) is reduced distally with a concomitant reduction in the metacone and metastylar lobe. However, in M4, unlike the preceding molars, the metaconule is larger than the paraconule. Numerous isolated lower teeth are referred to S. encinensis. These are all from localities or the same fossil horizons that yield upper teeth referable to S. encinensis, and so referral is not in doubt. In addition, a partial dentary with m1-2, NMMNH P-52885, is referred to this taxon. The dentary P-52885 preserves the alveolus for the double-rooted p3 mesial to the m1. A single mental foramen is situated below the mesial root of m1.
A single complete m4, 58982 (Figure 5.4-5.6) possesses a trigonid similar to that of m2-3. The talonid is narrower than the trigonid and relatively more elongate than in preceding molars. The entoconid is lower and smaller than the hypoconulid. Discussion. A small sample size available to study for both S. encinensis and S. johansoni hampers comparison. The holotype of S. johansoni (NMMNH P-59304), an M3 from locality L-7583, is over 14 percent smaller in buccolingual width than the holotype (21594) and a referred specimen (18587) of S. encinensis. A student T-Test of pooled m1-2 length (Table 2, Table 3) of Swaindelphys encinensis (n=4) and S. johansoni (n=2) results confirms that the difference between the two samples is statistically significant (P-Value = 0.0006). There is no overlap in tooth measurements between samples of these taxa. Swaindelphys encinensis is the largest metatherian reported from the late early Paleocene (Torrejonian) of western North America and is subequal in size to the Puercan taxon Thylacodon pusillus and T. cf. T. pusillus (e.g., Standhardt 1980, table 5; Williamson, unpubl. data; Archibald 1982, table 24; Clemens 2006, table 1). Johanson (Johanson 1996a) erected the genus Swaindelphys and described the species S. cifelli from Swain Quarry of northern Wyoming, noting that whereas the upper teeth possessed some characters typical of herpetotheriid metatherians such as a V-shaped centrocrista, the lower molars lacked some features typical of derived members of herpetotheriidae including such features such as a distally-directed hypoconulid that is located distal to the entoconulid. Johanson (1996a) further described the lower teeth of S. cifellii as being very similar in morphology, but smaller than those of Thylacodon pusillus or T. cf. T. pusillus described by Archibald (1982). The lower teeth of S. encinensis are similar in size to T. pusillus, but have lower, more robust trigonid cusps, a relatively lower, less trenchant entoconid, and an m4 entoconid that is smaller than the hypoconulid.
Family Peradectidae Crochet, 1979
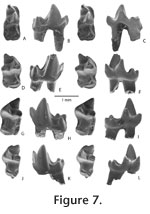
"Peradectes? n. sp. C" (Williamson and Lucas 1992), p. 120; (Williamson 1993), p. 102; (Williamson 1996), p. 34. "Peradectes? n. sp. D" Williamson and Lucas, 1992, p. 120; Williamson, 1993, p. 102; Williamson, 1996, p. 34 (in part). Holotype. NMMNH P-59512, right M3 from NMMNH locality L-7583. Referred specimens. From NMMNH locality L-312, NMMNH P-57803, left M2; 58826, left m1; 58846, right M1; 58849, left p3; 58854, right m1; 58927, left m2 or 3; 58933, right m1; 59014, right m4; 59015, left M1; 59062, left M3; 59094, left m4; 59195, partial right M3; 59228, partial right M1; and 59342, left m1. From NMMNH locality L-5652, NMMNH P-43089, left m1. From NMMNH locality L-6315, NMMNH P-53929, left m4. From NMMNH locality L-6398, NMMNH P-53881, right m1; 53936, partial right M2. From NMMNH locality L6898, NMMNH P-56974, right m2 or 3; 56976, partial left m2 or 3; 56977, right m2 or 3; 57819, right M3; and 57820, left M2. From NMMNH locality L-7583, NMMNH P-59050, right m1; 59059, partial right m2 or 3 (talonid); 59063, left M1; 59283, left m1; 59284, left m1; 59288, right M2; 59291, partial right m2 or 3; 59292, right partial M3; 59313, left M2; 59319, left M2; 59334, right m2 or 3; 59338, right m1; 59341, left m4; 59342, left m1; 59344, left m2 or 3; 59380, left m2 or 3; 59391, left m2 or 3; 59405, left m2 or 3; 59407, right m1; 59408, left m1; 59422, left m4; 59450, left m4; 59458, partial left M4; 59459, right M2; 59460, left m1; 59465, partial left M3; 59482, right m1; 59483, left M3; 59484, right M2; 59485, partial left M?; 59486, left partial dentary with m4; 59496, left m1; 59499, partial left M2 or 3; 59507, left m1; 59508, right M1; 59509, right M2; 59537, partial left M3; 59541, right m2 or 3; 59542, right m2 or 3; and 59545, left m2 or 3. Horizon and locality. From several localities of the Nacimiento Formation, San Juan Basin, New Mexico, distributed within fossil zones included in both the Pantolambda carivictum – Mixodectes pungens and Mixodectes pungens zones (Williamson 1996) and considered to be middle through late Torrejonian (To2-3) in age (Lofgren et al. 2004, Figure 2). Etymology. Combines kopros, (Greek, dung) with exoche (Greek, point) after Coprolite Point (NMMNH locality L-6398) from which the first specimens were collected (Taylor 1984). Diagnosis. Smaller than P. elegans (Matthew and Granger 1921), P. pauli (Gazin 1956), and P. californicus (Stock 1936); differs from all species of Peradectes including P. minor by more expanded and buccally more strongly convex metastylar lobe, deeper ectoflexus, relatively larger stylar cusp C, larger and more distinct conules and internal conular wings; further differs from P. minor by possessing a relatively more elongate m4 trigonid and narrower m4 talonid. Description. Peradectes coprexeches is represented by numerous specimens, all consisting of isolated teeth or small jaw fragments with single teeth. M1 is triangular in occlusal view with a stylar shelf that narrows mesially. A triangular parastylar lobe projects mesial to the paracone. The buccal margin of the tooth is nearly straight with a shallow ectoflexus distal to stylar cusp B. Stylar cusp B is the highest of the stylar cusps, followed by stylar cusps D, C, and A. Stylar cusp C is present as an elongated cusp along the margin of the stylar shelf in most specimens, but is represented as two smaller, closely appressed cusps in one specimen (59063; Figure 6.2), similar to what Clemens (2006) described for P. minor. The metacone is higher and longer than the paracone. The centracrista is straight. The preparacrista is short and extends buccally to the mesial base of stylar cusp B. The conules are subequal in size, and the internal conular wings are small, but distinct The postmetaconular crista descends to near the distolingual base of the metacone. M2 is more transverse than M1 with a wider stylar shelf buccal to the paracone. The ectocingulum of the metastylar lobe expands buccally in a convex arc, and the three mesial stylar cusps, A, B, and C form a nearly straight line that is directed nearly mesially. M3 differs from M2 in having a more buccally expanded parastylar lobe and a relatively deeper ectoflexus. Both are buccally rounded in occlusal view and the ectoflexus is approximately centered on stylar cusp C. In some specimens (e.g., 57819 and 59062; Figure. 6.5, 6.6, respectively), a deep notch is present, in occlusal view, immediately distal to stylar cusp C. However, in one specimen (59512; Figure 6.4) no such notch is present. A partial M4 (Figure 6.10) possesses a reduced metastylar lobe, lacks a distinct stylar cusp C, and has a shallow ectoflexus. Lower teeth are represented by a single p3 and numerous lower molars. The p3 closely resembles that of the holotype of P. elegans (AMNH 17376). The principle cusp is high with a convex mesial border. Distally, a crest descends to a narrow talonid, which supports a single blade-like median cusp. Mesially, a crest descends and curves lingually to the base of the crown where it merges with a lingual cingulid. In buccal view, the face of the tooth is convex.
The m2-3 (Figure 7.4-7.9) differs from m1 in having relatively shorter and wider trigonids with a paraconid that is positioned near the mesiolingual corner of the tooth. The entoconid is relatively lower and shorter so that it is subequal in size to the hypoconulid or smaller. It is conular rather than buccolingually compressed. The m4 (Figure 7.10-7.12) is smaller than m2 or 3, but with a relatively more elongate protoconid. The talonid is narrower than the trigonid, but as in the m2 or 3, the entoconid and hypoconulid are subequal. However, the postcingulid is relatively weaker and an ectocingulid is lacking from the ectoflexid. Discussion. All specimens of Peradectes coprexeches were recovered using screenwashing methods. This material comes from several localities, and most specimens are from a single locality, L-7583. Three specimens, NMMNH P-59508, 59509, and 59512, representing an M1, M2, and M3, respectively, were recovered from the same small batch of screenwashing matrix and likely come from a single individual. Peradectes coprexeches differs other species of Peradectes by its more expanded metastylar lobe, deeper ectoflexus, and relatively larger stylar cusp C. The buccal margin of the metastylar lobe is strongly convex in occlusal view and expands buccally beyond the parastylar lobe, whereas the metastylar lobe of P. elegans is relatively flat and does not extend buccally significantly beyond the parastylar lobe. Taylor (1984) reported an unpublished metatherian taxon, "Peradectes coprexeches," from a single locality (NMMNH locality L-6398; "Coprolite Point") in Kutz Canyon of the northern San Juan Basin (Figure 1). Most of the specimens that he referred to this taxon were unavailable for this study. However, we refer one specimen available to us from this locality, a partial M2 (NMMNH P-53936; = UALP 14599, Table 1). It closely resembles M2s referred to this taxon recovered from other localities in the southern San Juan Basin in the distinctive shape of the buccal margin of the tooth, with a relatively pronounced ectoflexus compared to other species of Peradectes, and the relatively large size of stylar cusp C. A fragment of a lower tooth from this locality (53881) is tentatively referred to this taxon. Taylor (1984) referred an additional specimen, a partial M1, (UALP 10498 from UALP locality 7671) to "P. coproxeches." This specimen was not available to study, but based on the measurements reported for this specimen (Taylor 1984, table 5; width = 2.1), it falls outside of the size range for this taxon. Based on the measurement, it might be referable to Swaindelphys johansoni (above). A single lower tooth, an m1 (NMMNH P-43089) from locality L-1870, is also tentatively referred to this Peradectes coprexeches. It is similar in size and morphology to m1s referred to Peradectes coprexeches recovered from other localities reported here. |
|