|
|
|
SYSTEMATIC PALEONTOLOGY
Family Cricetidae
Fischer, 1817
Genus MIMOMYS Forsyth Major, 1902
Mimomys hordijki
van Kolfschoten
and Tesakov, 1998 1998 Mimomys hordijki sp. nov.; van Kolfschoten and Tesakov, p. 187–192, figs. 1, 2.
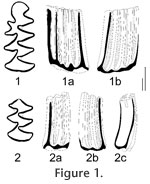
Type. Lower m1 (2.8 x 1.22 mm) and m2 (1.74 x 1.05) originating from the same mandible, BRM (Brielle Local Museum), no. Z3-201 (Figure 1). Material. 12 complete lower m1s (Z7-1, Z11-2, Z11-1, Z6-1, Z7-2, Z7-3, Z10-1, Z2-17, Z2-2, Z2-166, Z2-1, Z5-167) except the holotype, and one damaged m1 without PL (Z9-1), 2 M3 (Z6-2, Z9-2). All specimens are in collection of Brielle Local Museum (BRM), The Netherlands: ZU-4 (=Zuurland Unit 4, depth level of 61 to 66 m), Early Pleistocene, Gelasian, Late Villanyian, MN17.
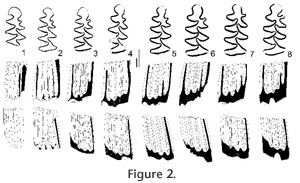
Lower m1. The anteroconid has a strongly developed Mimomys-ridge opposing the posterior part of T5. Reentrants of the anteroconid are deep; T4 tends to be more isolated in advanced stages of wear. The mimosinuid is normally high (e.g., Figure 2: 2) or of medium height (Figure 2: 5); it is only rarely not developed (Figure 2: 4). The posterolingual tract (HSLD) is slightly higher than the labial one. The base of the islet reentrant (BRA3) is deep. No signs of an enamel islet are present, not even in the two juvenile specimens (Figure 2: 1, 2). Lower m2. The occlusal surface consists of a PL and two pairs of broadly fused triangles, T1–T2 and T3–T4 (Figure 1: 2). The base of the molar is strongly curved lingually indicating the pleurorhizal condition (at the outer side of the incisor). The base of LRA1 is deep, and the height is not reduced by the contact with the incisor. H = 3.50, ASD> 4.14, HSD = 3.33, HSLD = 4.00, Lbas = 2.00, HH-index = 5.20.
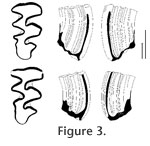
Comparison. The Hordijk's Mimomys differs from Mimomys monohani L. Martin, 1972 from Mullen l.f. (of possibly Irvingtonian age) in a somewhat larger size, smaller amount of cement and less divided occlusal elements in the m2. All these characters tentatively indicate a more advanced evolutionary stage of M. monohani. Mimomys virginianus Repenning and Grady, 1988, late Irvingtonian in age from Hamilton Cave, differs from M. hordijki in having smaller size, more abundant cement accumulations, a more anterior position of the Mimomys-ridge (opposite the center of T5), and more separated T2 and T3 in the M3. Some molars of M. virginianus from Porcupine Cave (Bell and Barnosky 2000, p. 101, figure 4: D) seem to have thicker leading edges of the enamel band (Microtus-type or positive differentiation sensu R. Martin 2003). Mimomys dakotaensis R. Martin, 1989, Java l.f. of Irvingtonian age, is similar to M. hordijki in the structure of the only known m1 (R. Martin, 1989, p. 443, figure 5: A; figure 4: 3). However, the m2 and the M3 have more separated occlusal fields than in the Dutch Mimomys hordijki vole.
Mimomys hordijki and North American Irvingtonian Mimomys show close morphological similarities. The most important distinctive features are the structure of the m1 with a strongly developed Mimomys-ridge and without enamel islets, and of an M3 with a hook-like posterior part and a deep postero-lingual re-entrant. This group is known from the Early Pleistocene (Gelasian, Villanyian LMA) of Europe, the Early Pleistocene (Gelasian to 'Calabrian', Kutuyakhian through Olyorian) of western Beringia (Kolyma lowland) and eastern Beringia (Repenning 2003), and from Early Pleistocene (Irvingtonian) faunas of conterminous United States. This indicates a high latitude Holarctic distribution of this group during the early Early Pleistocene and a dispersal to more southern regions of North America in the late Early Pleistocene. A proper name for this group of mimomyoid voles has yet to be established.
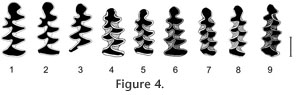
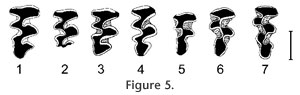
Evolutionary polarities of characters evolving in vole dentitions permit reconstruction of the following sequence of species attributable to Cromeromys. The most primitive of the forms described so far is Early Pleistocene M. hordijki from Europe. The sparse cement and still-measurable complete dentine tracts in younger individuals are the most reliable indicators of its evolutionary level. A more primitive form from the Kutuyakh beds (early Late Pliocene, ca. 2.5 Ma) of northeastern Siberia is still undescribed and only listed in the literature (Zazhigin 1997). Also undescribed are Early Pleistocene (Irvingtonian) records from eastern Beringia (Repenning 2003). The Irvingtonian forms from the conterminous United States show a number of characters that are more advanced than those of M. hordijki, including more abundant cement, smaller size, a more anteriorly shifted T6, and more separated triangles in the m2. Range and occurrence. The species is known from the type locality, Zuurland, The Netherlands, Zuurland faunal unit 4 (ZU-4), Late Villanian, MN17, Early Pleistocene. Recently, we identified a fragment of an m1 of this species in the broadly synchronous Dutch locality, Tegelen-Maalbeek (so far unnumbered specimen, collection of Faculty of Archaeology, Leiden University, The Netherlands). The Taxonomic Position of LemmiscusStudying the material referred to Mimomys ("Cromeromys") hordijki led to the discovery of a number of similarities between this species and species referred to the genus Lemmiscus. Those similarities suggested that it was worth exploring the possibility that the two are related. The Nearctic sagebrush voles of the genus Lemmiscus have long been suspected to be related to Palearctic lagurines or steppe lemmings (genera Lagurus and Eolagurus) because of obvious similarities in ecology and morphology of these voles (Gromov and Polyakov 1977; Carrol and Genoways 1980), and for a long time both were assigned to the same genus Lagurus. However, karyological data support close sister-group relationships of Lemmiscus with Arvicolini (Modi 1996) and most recent reviews reestablished the independent generic status for the North American Lemmiscus (Carleton and Musser 2005). A grouping of Lemmiscus with Arvicolini is supported by a number of morphological features. One feature includes external molar cement shared by both Lemmiscus and Microtus s.l. but absent in all lagurines. Views on the taxonomic value of this character range from a total disregard to the understanding of its utmost importance (Zazhigin 1980). Another tribe-specific character of Lagurini, which is lacking in Lemmiscus, is the so-called lagurine foramen (Zazhigin 1980) or foramen lagurinorum, the small opening for a nerve or blood vessel on the mesiodorsal surface of the mandible, below the antero-internal part of first lower molar. A conspicuous dental character shared by Lemmiscus and Lagurini is the so-called Lagurus fold in the lingual reentrants of the upper molars (R. Martin 1989). However, Lemmicus has a distinct pattern of these indentations including their presence in posterointernal reentrant angles of the M1 and M2, a character never observed in lagurines. Moreover, additional enamel folds frequently occur in molars of different groups of rootless voles, including, for example, Arvicola, Microtus, and Alticola. Thus, these characters likely originated in parallel in many lineages as an apparent way to strengthen the molar structure, rather than representing remnants of ancestral cuspate elements (Martin 1989) such as a 'protoconule' (Hinton 1926). Another important dental feature common to both Lemmiscus and Lagurus is the additional dentine tract in the lower m1. In the evolution of the dentition of voles, the dentine tracts originated in primitive rhizodont forms from undulations of the enamel-dentine boundary. With increasing hypsodonty, dentine tracts also grow, serving as anchoring zones of molars in the alveoli (L. Martin 1979; Rabeder 1981). After the evolutionary acquisition of rootlessness in molars, dentine tracts were transformed into permanent interruptions of the enamel wall present in molars for most of the animal's life. Only in unerupted or hardly worn teeth of juvenile rootless voles can the uninterrupted tips of dentine tracts still be observed. The number and positions of the dentine tracts are stable in molars; they are regarded as an important taxonomic character. Normally, many arvicoline lineages have only three basic dentine tracts in the first lower molars: two at both sides of the posterior loop and one at the anterolabial side of the anteroconid. The first lower molars of Lemmiscus and Lagurus both have, apart from the normal arvicoline set of tracts, an additional tract along the BRA4 or T6. This tract topologically and homologically corresponds to the Mimomys-ridge, a feature that is particularly well-developed in Eurasian voles of the genera Mimomys and Borsodia. This ridge often bears an additional dentine tract (mimosinuid, according to the terminology of Rabeder); the development of that tract is variable. However, mimomyoid features, including the ridge and the corresponding tract, are usually reduced in most lineages with an increasing hypsodonty of the molars. None of almost 60 extant species of Microtus show additional tracts in the T6. The stable presence of this character in Lemmiscus and Lagurus implies a strong development of the Mimomys-ridge and its tract in their rhizodont ancestors. The history and origin of the additional tract in Lagurus is relatively well known and can be traced back in time at least to the Early Pleistocene (Biharian LMA) and eventually to early Early Pleistocene (Villanyian) species of the ancestral genus Borsodia that had both the ridge and tract (Zazhigin 1970, 1980; Rabeder 1981; Tesakov 1993, 2004). The history of the additional dentine tract in Lemmiscus is yet to be traced back in time. In many published accounts, interruptions of the enamel band are frequently not studied and not illustrated. Anyway, though quite rare, this feature cannot automatically be considered as a synapomorphy. Despite of the similarities between Lemmiscus and Lagurus, taking into account the different arguments mentioned above, it can be stated that a close affinity between Lemmiscus and the Palearctic Lagurini should be rejected and most morphological similarities of the two groups may be interpreted as a parallel evolution in conditions of open, steppe-like landscapes. Looking for Possible Ancestors of LemmiscusThe rejection of an affinity with lagurines and the strong evidence for microtine affinities of Lemmiscus provides a clue for the search of an ancestral group for Lemmiscus. In fact, Repenning (1992) already assumed that Lemmiscus originated from the group of primitive Allophaiomys voles, a presumably ancestral group for modern Microtus. Allophaiomys is hypothesized to have evolved in Eurasia from a group of hypsodont Mimomys (Tcharinomys) species (M. gr. tornensis-haplodentatus) with abundant external cement and a strongly reduced complex of primitive mimomyoid characters such as a Mimomys-ridge and enamel islets (Rabeder 1986; Tesakov 1998, 2004). Lemmiscus is distinct from all the hypsodont Mimomys (Tcharinomys) species and all descendant Microtus species in the stable presence of an additional dentine tract in the m1. On the other hand, a close morphological and genetic similarity to Arvicolini permits the suggestion of a hypothetical Mimomys ancestor for the modern sagebrush voles. To fit this hypothesis, the ancestral form should display a strongly developed mimosinuid and a dentine tract homological to the additional tract on the sixth dental prism of the m1 of Lemmiscus. Although several Eurasian Mimomys clades show a developed mimosinuid (Rabeder 1981; Zheng and Li 1986; Tesakov 2004), it is rarely so well-developed as in the Early Pleistocene (Villanyan) Mimomys hordijki from The Netherlands and in Mimomys (Cromeromys) spp. in the Irvingtonian faunas of North America. This fact, combined with the occurrence of the latest Mimomys (Cromeromys) and earliest Lemmiscus in the same Nearctic region, suggests a phyletic link between Mimomys hordijki and the genus Lemmiscus. Fossil Record and Morphology of Lemmiscus: Testing the Cromeromys OriginThe known history of Lemmiscus starts in the late Early Pleistocene with the record from the SAM Cave in New Mexico (Repenning 1992; Rogers et al. 2000). The next oldest record is from the sequence of Porcupine Cave in Colorado (Bell and Barnosky 2000; Barnosky and Bell 2003; Bell et al. 2004) ranging in age from latest Early Pleistocene to the Middle Pleistocene. Contrary to the common dental pattern of the m1 with five or more closed triangles in modern Lemmiscus, the fossil record shows an increasing number of more primitive four-triangle morphotypes back in time (Barnosky and Bell 2003). A coining of a separate chronotaxon name is recommended for samples with the predominance of this morphology. The short co-occurrence or time overlap of presumed ancestral (rhizodont) and descendant (rootless) forms in Late Irvingtonian (Bell and Barnosky 2000) do not contradict the hypothesis of a Mimomys–Lemmiscus phyletic lineage. In fact, it could be the time of a rapid disappearance of molar roots characterized by the occurrence of transitional populations with rooted and rootless morphotypes. The known transitions from rhizodont to arhizodont condition in voles are usually so fast in time that transitional populations are only rarely represented in the fossil record (Tesakov 1993, 2004; Maul et al. 2000). A hypothetical morphological transition of the m1 morphology from Mimomys (Cromeromys) to Lemmiscus conditions constructed with published examples of dental structure and with regard to geological age is presented in Figure 4. Note the gradual anterior shift of the Mimomys-ridge and the successive separation of the dental triangles. If valid, this sequence shows the formation of the sixth dental triangle of the modern Lemmiscus from the gradually enlarged Mimomys-ridge. The strongly developed mimosinuid of Mimomys (Cromeromys) gave rise to the enamel band interruption at the tip of T6 in Lemmiscus. A similar reconstruction of the transition in morphology of the upper M3 is presented in Figure 5. Note the close morphology of Mimomys species (Figure 5: 1–4) and considerable change in the shape of the M3 of the rootless Lemmiscus (Figure 5: 5–7). However, such a character as the confluence of AL and T2 can be traced from the Mimomys stage to the early Lemmiscus (Figure 5: 5). According to Barnosky and Bell (2003), the predominant morphotype of the M3 with a shallow antero-labial re-entrant is characteristic for the Early Pleistocene Lemmiscus, and it is completely replaced by a morphotype with deep reentrant in the modern species. Noteworthy is a lack of apparent complication in M3 in rootless Lemmiscus parallel to a trend to more complex morphology (addition of dental triangles) in m1s. But given the much smaller amount of material on M3s any straightforward conclusions would be premature at the present state of knowledge. |
|