|
|
|
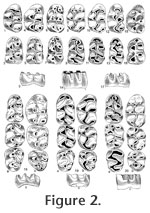
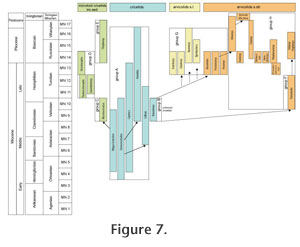
INTRODUCTIONArvicolids are muroid rodents that first appeared in the Late Miocene of Eurasia and North America and have been widely dispersed in the Northern Hemisphere since that time (Repenning 1987; Repenning et al. 1990; Fejfar et al. 1997). Today, arvicolids include the lemmings, muskrats, meadow mice, red-backed voles, heather voles, sagebrush voles, and the mole voles. They are one of the most successful groups of rodents to inhabit the northern continents during the late Cenozoic. The fossil record of arvicolids is outstandingly rich. Their high-crowned prismatic molars can be readily distinguished from those of any other rodent group. Arvicolid cheek teeth and other skeletal remains can easily be obtained in great quantities by washing and sieving sediment. Enormous numbers of sites scattered widely in Eurasia and North America have produced diverse assemblages of fossil arvicolids. An increasing body of literature provides much information about their taxonomy, distribution, biostratigraphy, evolution, and phylogeny. However, data about the roots and early history of arvicolids are rather limited (e.g.,Hinton 1926; Repenning 1968; Kretzoi 1969; Fejfar and Heinrich 1983; Nesin and Topachevskij 1991; Topachevskij and Nesin 1992; Fejfar et al. 1997; Chaline et al. 1999; Agadjanjan 2009) and a generally accepted model about the origin of voles and lemmings has not yet been established. Today, there is no doubt that arvicolids evolved from a specialised branch of advanced cricetid rodents designated by the informal name "microtoid cricetids" by Schaub (1934). "Microtoid" means arvicolid-like, since not all of these forms evolved to the group of true arvicolids. The terms arvicolids and microtids (Arvicolidae and Microtidae) are currently used as synonyms, traceable back to Miller (1896) who considered the genus Arvicola as a junior synonym of Microtus and therefore rejected the name Arvicolidae Gray, 1821 and used Microtidae Cope, 1891 (Kretzoi 1990). The systematic rank of arvicolids has been debated for many years (e.g., Kretzoi 1969; Musser and Carleton 1993 and references herein). Many authors refer arvicolids to subfamily rank (Arvicolinae), but given high species diversity and the specialised adaptive level, they are considered by some workers to be of family rank (e.g., Niethammer 1982), a view shared by the present authors. Extant arvicolids and cricetids are morphologically well distinguished, but fossil forms display transitions in many characters. Arvicolids are ecologically and morphologically adapted to digging and eating less nutritious vegetative plant remains, mainly monocots (Gramineae), and do not hibernate, whereas Eurasian cricetids mainly eat seeds and roots and do hibernate (Niethammer 1982). A crucial feature of arvicolid molars is that they are, or show tendency to become, hypsodont. Parallel and angled lateral walls (in some older forms slightly conical) cause a more or less constant occlusal surface in all grinding levels, which is called a "prismatic" form. The chewing surface is flat with serial prisms alternating on buccal and lingual sides. The enamel band is undifferentiated in primitive taxa and mainly differentiated in derived taxa. Cricetids originally have brachyodont and bunodont molars with cutting edges at the tubercles and not planar occlusal surface. Arvicolids and cricetids differ in several cranial features and in enamel microstructure, adapted to various modes of chewing and biomechanical forces resulting from the positions of muscles (Repenning 1968; von Koenigswald 1980). However, since it is evident that arvicolid-like characters evolved independently within cricetids at several times, the question remains as to which cricetid rodents gave rise to "true" early arvicolids. In the following, we summarise data from the relevant time interval and discuss possible phylogenetic relationships between the various microtoid cricetids and early arvicolids in order to enlighten the basal phase of arvicolid evolution. Primitive cricetids and arvicolids of Eurasia are the main focus of the present paper. The biochronological framework used in the present paper is that of Repenning (1987), Tedford et al. (1987, 2004), Fejfar and Heinrich (1990), Fejfar et al. (1997, 1998) and Martin (2003a). The use of MN zones of the European Neogene mammal chronology follows Mein (1990, 1999), usage of the Neogene mammal faunal units (NMU) in China Qiu et al. (1999) and Qiu and Li (2003). A graphical review of the significant taxa of cricetids, microtoid cricetids and early arvicolids is given in Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, and Figure 6. The palaeogeographical and stratigraphical ranges of the considered taxa are shown in Figure 7. In the following descriptions, lower case denotes lower teeth and upper case denotes upper teeth. For elements of the occlusal surface the terminology of Fahlbusch (1964) and van der Meulen (1973) is used.
|
||||||||||