|
|
|
|
:
BASKIN |
|
SYSTEMATIC PALEONTOLOGY
Family MUSTELIDAE
Fischer de Waldheim, 1817 Diagnosis (From Baskin 1998, p. 164). "Small mustelines with a deep jaw, crowded premolars, and p1 absent; P4 primitively with a small anterior protocone and narrow lingual cingular shelf, that is more expanded in derived forms; M1 with an expanded hypocone and a more separate protocone-metacone; and m1 with an open bladelike trigonid, a lingually expanded and basined talonid with a posterolingual cingulum extending from the base of the reduced metaconid to the hypoconid, and hypoconid separated from the protoconid by a small notch." Included genera. New World: Galictis, Stipanicicia, Trigonictis, Sminthosinis, Lutravus, Cernictis, and possibly Lyncodon; Old World: Enhydrictis, Pannonictis, Oriensictis, and possibly Trochictis. Discussion. Pocock (1921) established the Grisoninae as a subfamily of Recent Mustelidae for the Neotropical Grison (=Galictis vittata) and Grisonella (=Galictis cuja), the greater and lesser grison. Pilgrim (1933) added the fossil European genera Trochictis, Enhydrictis, and Pannonictis; the late Miocene "Mustelide gen. indet. sp. n." from China (Zdansky 1927; Ficcarelli and Torre 1967); and the Neotropical Pleistocene and Recent Tayra (=Eira) to the Grisoninae. Subsequently, Schreuder (1935) excluded both Eira and Trochictis from the Grisoninae, because of the characteristics of their upper carnassials. Furlong (1932) described Lutravus halli as a new genus and species of otter (Lutrinae). Gazin (1934) added two species to Lutravus (?), based on two lower jaws from Idaho. Although he described the three species as otter-like, Gazin (1934) noted the many similarities of the lower dentitions of Lutravus to the grisonines Enhydrictis, Pannonictis, and Galictis. After recovering upper carnassials of one of his two species, Gazin (1937) transferred the Idaho species to Canimartes?, a genus known only from a few isolated teeth and tooth fragments (Baskin 1998). Repenning (1967) suggested that Gazin's (1934) two species could be derived from the European Trochictis. Zakrzewski (1967) assigned the two species to Trigonictis (Hibbard 1941), which had been recognized as a galictine by Reig (1956), but left Furlong's species as the sole member of Lutravus. Reig (1956) emended the subfamily name to Galictinae and added the North American Neogene Lutravus and Trigonictis, and two Neotropical genera, Lyncodon (the extant Patagonian weasel) and the Pleistocene Stipanicicia. Reig (1957) placed Lutravus in the Galictinae because of the grison-like features, especially of the lower dentition as noted by Gazin (1934). Reig (1957) also included Cernictis, the subject of the present paper. Bjork (1970) included Trochictis, Enhydrictis, Trigonictis, Sminthosinis, Galictis, and Grisonella in the Galictinae. The systematic positions of Trochictis and Eira have been debated. The early to middle Miocene (MN4-MN5) Trochicits has been considered the most primitive galictine (e.g., Pilgrim 1933; Ficcarelli and Torre 1967; Bjork 1970). More recently, it has been assigned to the Melinae, the Old World badgers (e.g., Petter 1971; Ginsburg and Morales 2000), although some still treat it as a galictine (e.g., Ogino and Otsuka 2009). Pilgrim (1933) concluded that Eira retained primitive characters of the skull and P4 similar to Trochictis and therefore was a galictine. Reig (1957) agreed with this reference, but Ficcarelli and Torre (1967) excluded Eira from the Galictinae, because of differences in the dentition and auditory bulla from other members of the subfamily. As suggested by the upper cheek teeth (Baskin 1998) and confirmed by molecular phylogeny (Fulton and Strobeck 2006), Eira is not a galictine, but is more closely related to Martes. In Eira, the P4 has a well developed protocone on a spur opposite the paracone and lacks a lingual shelf. Molecular phylogeny not only supports the subfamilial status of Galictis, but also indicates that the Old World Ictonychini (zorillas, etc.) are galictines (Fulton and Strobeck 2006; Koepfli et al. 2008). Other recent discussions of the contents and characteristics of fossil and recent Galictini can be found in Ray and others (1981), Baskin (1998), Morlo and Kundrát (2001), and Ogino and Otsuka (2008). Genus CERNICTIS Hall, 1935 Type species. Cernictis hesperus Hall, 1935. Type and only specimen. UCMP 22968, left mandible with p4-m1. Type locality. Pinole Junction, Site 1, Pinole Tuff Formation, Contra Costa County, California. Late Hemphillian North American Land Mammal Age Included species. Cernictis hesperus, C. repenningi n. sp. Emended diagnosis. There are three crowded lower premolars. The p2 is two rooted. The p4 possesses a posterior accessory cusp. The m1 is relatively short relative to the p4; the talonid is less than half the length of the trigonid; the talonid is semi-basined, and the metaconid is moderately developed. Comparisons. The deep jaw, crowded premolars, and internally expanded talonid with a posterolingual cingulum extending from the base of the reduced metaconid to the hypoconid on m1 are diagnostic of the galictin affinities of Cernictis. Cernictis differs from Trochictis in loss of p1, having a more crowded dentition, and having the mandible deep, especially anteriorly. It differs from Galictis, Enhydrictis, Pannonictis, Trigonictis, Sminthosinis, Lutravus, and Oriensictis in possessing a posterior cusp on the p4 and in having a shorter and less well basined talonid on the m1. It differs from Galictis and Eira in having a two rooted p2, and a larger metaconid, more trenchant hypoconid, and more basined talonid on the m1. Eira also lacks a posterior cusp on the p4. Hall (1935) described the lower jaw of his new genus and species, Cernictis hesperus, as having some similarities to marten-like forms such as Martes or Gulo, but not being closely related to either. Reig (1957) concluded that the p4 and m1 of Cernictis were most similar to those of Eira, and included both in the Galictinae. In contrast to other galictins, Cernictis, possesses a relatively short talonid on m1, a feature also present in Eira. Characteristics of the upper dentition, especially the P4, have been important for assigning genera to the Galictini. Enhydrictis, Pannonictis, Trigonictis, and Galictis are characterized by a P4 with a triangular occlusal outline, a lingual shelf and a small protocone. Sminthosinis (Bjork 1970) has P4 with a relatively large protocone and unexpanded lingual cingulum. Undescribed material from the late Miocene (Turolian) of China in the collections of the AMNH adds support that Cernictis is a primitive galictin, because the upper dentition is otherwise unknown in Cernictis. Information with a skull and lower jaw indicate it is from the Baodean (MN 12, ca 8 Ma) locality Ma Chia Liang Kou in northern Shanxi Province. The lower jaw and dentition have characteristics of Cernictis, but with more slender premolars and a better developed metaconid on m1. The P4 has a small anteriorly placed protocone and a very weakly developed internal cingulum. The M1 is not expanded internally. The lower dentition resembles the figures of Baranogale adroveri from the Turolian (MN12) of Spain (Petter 1964). Baranogale is an extinct member of the Ictonychini. Spassov (2001) questioned the assignment of Petter's species to this genus.
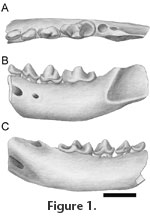
Cernictis repenningi n. sp.
Referred material. UALP wc 251/6, right M1 fragment; 260/6, m2. Distribution. White Cone (UALP locality 6). Bidahochi Formation, northeastern Arizona; Hemphillian North American Land Mammal Age. Etymology. Named for C.A. Repenning, collector of the holotype. Diagnosis. A species smaller than Cernictis hesperus Hall. There are three laterally compressed, crested, and crowded premolars. The p4 is relatively wide posteriorly. The protoconid of the m1 is only slightly higher than the paraconid and is lower than the main cusp of p4. The entoconid is absent. Description. There is no alveolus for a p1, between the p2 and the alveolus for the c1. The p2 is set obliquely in the jaw and has a single anteriorly directed cusp. A slight crest extends anteroposteriorly along the midline of the tooth, terminating in a small cuspule. The p3 has a single anteriorly directed cusp. There is a narrow anterior cingulum and a broader posterior cingulum. The anterior-posterior crest terminates in a small cuspule. On p4, the posterior accessory cusp is located slightly labial to the midline of the tooth. The anterior cingulid is low and very short and is connected to the wide and low posterior cingulid by a narrow external cingulid. The m1 is a relatively narrow, blade-like tooth. The protoconid is the tallest and most prominent cusp, with a weak antero-external cingulid. The apex of the paraconid is situated on the midline of the tooth. The metaconid is a robust cup situated on the posterior margin of the protoconid and connected to it by a weak ridge. It is nearly equal in height to the paraconid. Internally, the valley between the paraconid metaconid is open and indented at its base. On the external side, a narrow notch separates the posterior margin of the protoconid from the hypoconid. The talonid is short, semibasined, and slightly expanded internal to the metaconid. The hypoconid is trenchant and internal to the protoconid. A low posterointernal crest runs from the hypoconid to the base of the metaconid, closing the basin. The mandible is deep, especially anteriorly. There are two mental foramina; one below the p2, the other below a point between p3 and p4. The external half of a worn M1 and an isolated m2 are tentatively assigned to C. repennningi. The M1 has a metacone slightly smaller than the paracone. The two cusps are joined by a short loph. The m2 is small (L = 2.9, W = 3.0), single rooted, and circular in occlusal outline. The tooth is divided into anterior and posterior basins, plus a labial shelf. There is a low lingual cusp and a smaller labial cusp. Comparisons. Cernictis hesperus Hall (1935) from the Hemphillian Pinole Formation of California is known from a mandibular fragment with p4 and m1. It is larger than C. repenningi (Table 1). The p4 is much narrower posteriorly and has a better developed anterior cingulid cusp. The m1 is shorter relative to the p4 and has a smaller metaconid. The internal wall of the talonid is better developed than in C. repenningi. In C. hesperus, the protoconid of m1 is much higher than the paraconid of m1 and slightly taller than the main cusp of p4. In C. repenningi, the protoconid of m1 is somewhat higher than the paraconid of m1 and lower than the main cusp of p4. Lutravus halli (Furlong 1932) is much larger than Cernictis repenningi. The metaconid of the m1, as illustrated, appears to be smaller, more separated from the protoconid, and situated more posterior to the protoconid than in Cernictis. The mandible is not as deep relative to the cheek teeth as in Cernictis. Remark. Repenning et al. (1958) noted the presence of a "marten-like animal" from the Bidahochi Formation at White Cone Peak. Because the Bidahochi specimens were described in my unpublished Master's thesis (Baskin 1975) as belonging to Martes (Plionictis), this misidentification was repeated in subsequent publications (e.g., Lindsay and Tessman 1974; Lindsay et al. 1984; Morgan and White 2005; Hodnett 2010). Correct attribution to Cernictis was first published by Baskin (1998) and used in a faunal list of the Bidahochi Formation by Morgan and White (2005). Discussion. Immigration events are used as biostratigraphic markers to delineate and subdivide North American Land Mammal Ages (Tedford et al. 1987). The late Miocene in particular was marked by dispersal of carnivorans, including galictins, across the Bering Strait into North America (Repenning 1967; Qiu 2003). Trochictis has been considered the earliest galictine, making galictines distinct from other mustelids since perhaps the early Miocene (Pilgrim 1933; Reig 1957). If Trochictis is a meline, the earliest galictine may be Cernictis sp. from China, discussed above and dated at about 8 Ma. However, an occurrence of ?Lutravus from the late Miocene (ca 8 Ma) in the Ukraine is cited in Vangengeim and Tesakov (2008). In the New World, the earliest galictines are Lutravus halli and Cernictis repenningi, at about 8 Ma and 6.5 Ma, respectively. |
|