|
|
|
SYSTEMATIC PALEONTOLOGY
Order LAGOMORPHA Holotype. GSP 50951, right p3 from locality Y921, upper part of the Dhok Pathan Formation. Referred Material. GSP 50953, left P3 or P4; GSP 34504, left p3; GSP 34505, right dp3; GSP 34506, left dp4; GSP 34508 and GSP 50952, likely m1s; GSP 52500, left p4-m2; GSP 39389, right mX. GSP 53241, right M1 or M2 is tentatively referred. Occurrence. GSP 34504 - 34506, 34508 are from DP13. GSP 39389 and 50951 - 50953 are from Y921, "Bunny Hill" (these specimens are comparable in size and may represent the same individual); GSP 52500 is from Y456; and GSP 53241 is from Y975. All are from the upper part of the Dhok Pathan Formation, Siwalik Group, Potwar Plateau, Pakistan. Age. Y921, 7.35 Ma; Y456, 7.2 Ma; Y975, 7.2 Ma; DP13, 6.5 Ma. Etymology. Specific name "elongatus" is Latin for "prolonged," referring to the anteroposteriorly elongate trigonid on p3. Diagnosis. Lower third premolar roughly triangular in shape with anterior end elongated; connection between trigonid and talonid slightly oblique and located slightly labial to the midline. PER, PIR, and AER all distinct (primitive); AIR may be distinct or weak; AR lacking (primitive). Both PER and PIR extend approximately halfway across the tooth, but PIR is slightly deeper. AER and AIR anteroposteriorly elongate. TH smooth in PER, smooth to heavily crenulated in PIR; TN lightly to heavily crenulated. Enamel in AER lightly crenulated; smooth to lightly crenulated in AIR.
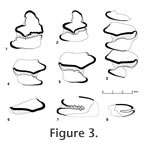
The P3 or P4 (GSP 50953; Figure 3.7) is incomplete: it is missing its external and much of its posterior portions. The internal reentrant (= hypostria) extends approximately two-thirds the way across the tooth and is heavily crenulated. The talon is wider, and likely slightly shorter, than the trigon. The M1 or M2 (GSP 53241, Figure 3.8) is likely from a sub-adult specimen because of its smaller size, possession of a relatively shallow internal reentrant with simple crenulations, and in side view, the size of the tooth increases toward its root. The internal reentrant of this specimen extends less than two-thirds the way across the tooth and is lightly crenulated. The talon is wider, and slightly shorter, than the trigon. GSP 53241 is only tentatively assigned to A. elongatus because the tooth is a sub-adult, from a relatively undiagnostic tooth position, and is the only lagomorph from locality Y975.
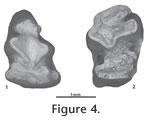
The dp4 (GSP 34506; Figure 4.2) has a separate trigonid and talonid. On the trigonid there is a gentle anteroexternal reentrant and a distinct deep internal reentrant (not observed on any adult post-p3 teeth). There is a single, wide crenulation on the anterior end of the talonid. The trigonid is wider than the talonid. The p3 (Figure 3.1, 3.2) is roughly triangular in occlusal outline. The trigonid is trefoil in shape with an elongated anterior end. A line drawn transversely through the external and internal extensions of the trigonid is slightly oblique, with the external side slightly inferior. There are four reentrants. The PER and PIR are deep, and each extends about halfway across the tooth, although the PIR on GSP 50951 is slightly deeper than the PER. These two reentrants are separated by the narrow, slightly oblique connection of (primarily) enamel between the trigonid and talonid. The PIR is almost closed off on its internal side. Within the PER and PIR, TN is lightly crenulated on GSP 34504 and heavily crenulated in GSP 50951. The TH is smooth within both the PER and PIR on GSP 34504. In GSP 50951 TH is smooth in the PER, but heavily crenulated in the PIR. The large AER is anteroposteriorly elongate, has cement extending continuously down the tooth, and forms almost a right angle with the long axis of the tooth. The AIR is distinctive on GSP 34504 (although not as large as the AER), is anteroposteriorly elongate, and forms almost a right angle with the long axis of the tooth. The AIR is weak on GSP 50951. The inside borders of both the AER and the AIR on GSP 34504 are lightly crenulated. AR is lacking. GSP 52500 (Figure 3.3) is an incomplete tooth row: the p4 is represented by only the external and posterior portions of the talonid, m1 is complete, and the m2 is complete except for the internal-posterior part of the talonid. GSP 34508 (Figure 3.4) and 50952 (Figure 3.5) are right m1s. These teeth have a wider trigonid than talonid. There is a distinct anteroexternal reentrant on the trigonid of GSP 34508 and 50952. This reentrant is lacking on the m1 and m2 of GSP 52500. GSP 39389 (Figure 3.6) includes only the posterior portion of the trigonid. The specimen does not appear to have an anteroexternal reentrant. Discussion. The genus Alilepus was erected by Dice (1931) based on the type species "Lepus" annectens Schlosser, 1924, from the late Miocene localities of Ertemte and Olan Chorea, Inner Mongolia, China (Schlosser 1924). Dice's (1929, p. 342) diagnosis (for Allolagus, but that name was pre-occupied, so changed to Alilepus in 1931) included a direct quote from Schlosser (1924) that the anterior lower premolar (p3) is formed "of two columns connected in the centre [sic] of the tooth by a bridge of dentine." White (1991) revised the genus based on study of Alilepus from the late Miocene to Pliocene of Eurasia and North America. White's (1991, p. 69) emended diagnosis was: "medium- to large-sized leporids with cranium and mandible fully modernized. P2 with deeply incised MAR and shallow EAR; p3 with PIR as deep or shallower than PER; PIR often pinched off to form an enamel lake; AIR shallower than PIR and usually missing; AR absent; TH smooth to slightly folded; AER shallow with smooth thin enamel." White (1991) noted that the distribution of the genus was the Turolian (late Miocene) to early Villafranchian (early Pliocene) of Eurasia and the late Clarendonian (late Miocene) to late Blancan (Pliocene) of North America. We assign the Pakistani specimens to the genus Alilepus based on the presence of a bridge connecting the trigonid and talonid and separating distinct PER and PIR, which are of about equal depth. The AIR is shallower than the PIR, and an AR is lacking. Several characters of the Pakistani specimens differ slightly from White's (1991) diagnosis: 1) the PIR of one of the two Pakistani p3s is slightly deeper than the PER; 2) the PIR is never pinched off to form an enamel lake, but there are only two Pakistani p3s; 3) the TH is smooth in both posterior reentrants on one Pakistani tooth, but on the other tooth it is smooth in the PER but well crenulated in the PIR, and 4) the AER has smooth to lightly crenulated thin enamel. The differences observed from White's (1991) diagnosis are not considered significant enough to warrant generic distinction. We conservatively assign the Pakistani specimens to one morphologically variable species. The geologically older p3 from Y921 (holotype, GSP 50951) is about 22% longer and 13% wider, has a shallower AIR, and generally has more crenulated enamel in the reentrants compared to the p3 from DP13 (GSP 34504). The m1 from Y921 (GSP 50952) differs from the p4-m2 from Y456 (GSP 52500), and the m1 from DP13 (GSP 34508) in having more crenulated enamel on the anterior end of the talonid. An anteroexternal reentrant is present on the m1s from Y921 and DP13, but not on the specimen from Y456. Comparisons with other species of Alilepus from Asia. Four generally accepted species of Alilepus are described from Asia: A. annectens Schlosser, 1924, A. longisinuosus Qiu and Han, 1986, A. lii, Jin, 2004, and A. laskarewi Khomenko, 1914. The type species is A. annectens ("Lepus" annectens of Schlosser 1924). Schlosser (1924) does not provide a formal diagnosis or detailed descriptions, so comparisons made here are with Qiu's (1987) more detailed descriptions (also lacking a diagnosis) based on additional collections from Ertemte and from Harr Obo.Qiu (1987) described the p3 of A. annectens as having a bridge connecting the trigonid and talonid, and possessing an AER, a PER, and a PIR (e.g., the AIR is lacking). The AER is wide and shallow, and the PER is narrow and deep, extending about halfway across the tooth. The PIR is more variably developed, but it never extends more than about one-third of the way across the tooth. Two out of about 14 p3s have an enamel lake instead of a PIR. Variable development leads to a variable bridge between the trigonid and talonid. Generally, toward the midline the PER and PIR both curve posteriorly. All folds persist to the base of the crown, and all are filled with cement. The dp3 of A. annectens is variable but similar in morphology to p3, except the PIR is deeper, and the bridge is narrower. The PIR may close off to form an enamel lake. When lightly worn, there is a shallow groove in the position of the AIR: this is lost rapidly in more worn specimens. The p3s of A. elongatus differ in having a distinct AIR and an AER, which is shallower, but more anteroposteriorly elongate than in A. annectens. The latter is likely related to the trigonid of A. elongatus being more anteroposteriorly elongate. Crenulations in the reentrants of A. annectens are minor or absent: there are moderate crenulations in three reentrants of GSP 34504, and strong crenulations in three reentrants of GSP 50951. Toward the midline, the PER and PIR do not curve posteriorly on GSP 50951. On GSP 34504 the PER is straight, and the PIR curves slightly posteriorly. On A. annectens the PER in particular curves strongly posteriorly. The dp3 of A. elongatus has a distinct AIR, versus its development only as a groove in lightly worn A. annectens. The p4-m2s of A. elongatus may have a distinct cement-filled anteroexternal reentrant; this reentrant is lacking in A. annectens. Alilepus longisinuosis Qiu and Han, 1986, is described from 69 specimens of latest Miocene age from Lufeng, Yunnan, China. This species is characterized by a variably developed bridge connecting the trigonid and talonid: the bridge may be wide, narrow, weak, or absent. The PIR is consistently deep, although it varies, and may be equal in depth to the PER or somewhat longer or shorter. Crenulations on the enamel of p3 are weak to absent. A distinct anteroexternal reentrant on p4 to m2 is lacking. The dp3 lacks an AER. The dp4 lacks an anterior lobe on the trigonid and has only two roots. The p3s of A. elongatus differ from those of A. longisinuosus with the former having a distinct narrow bridge between the trigonid and talonid, reentrants more crenulated, a distinct AIR, a more elongate trigonid, and a distinct anteroexternal reentrant on some p4 to m2s. The dp3 of A. elongatus has a distinct AER, and the dp4 has an anterior lobe on the trigonid. Alilepus lii, Jin, 2004, is known from a single mandible with p3-m1 from Huainan, Anhui, China. It is estimated as being latest Miocene in age (Jin, 2004). This species is characterized by its relatively small size, shallow AIR, strong crenulations on the TN of the PIR and the PER, rather wide bridge linking trigonid and talonid, and PIR distinctly less deep than the PER. On the p4-m1 there are distinct crenulations on the anterior side of the talonid, and an anteroexternal reentrant is lacking. The p3s of A. elongatus are similar to those of A. lii in general morphology, including having distinctive crenulations on the TN of the PER and the PIR. Alilepus elongatus has a better-developed AIR, somewhat more anteroposteriorly elongate trigonid, and a distinctly deeper PIR (resulting in a narrower bridge between the trigonid and talonid. The p4 and m1 of A. lii have distinct crenulations on the anterior side of the talonid, as do some specimens of A. elongatus. However, those specimens of A. elongatus with crenulations also have a distinct anteroexternal reentrant, a character lacking on A. lii. Among Asian species, A. lii is most similar to A. elongatus. Alilepus laskarewi Khomenko, 1914, was originally assigned to Lepus by Khomenko in 1914. It is from near the village of Taraklia in what is now Romania, and may be of Miocene age (Dice 1931). The name of the species (and its author) is sometimes translated from Russian into English as A. lascarevi Chomenko, 1914. Based on Khomenko's illustration (Khomenko 1914, plate 1.20) of the enamel pattern of p3, Dice re-assigned the species to Alilepus. Dice noted that A. laskarewi differed from A. annectens in the former's longer p3, with the greater length being primarily in the area anterior to the main pair of reentrant angles (likely the PER and the PIR). Dice (1931) also mentioned the presence of an AER on p3, and seemed to imply this reentrant was lacking on A. annectens (but it is present). The p3 of A. laskarewi includes an AER, a PER, and a PIR. The AR and AIR are lacking. The PER extends a little over halfway across the tooth and curves posteriorly near the median longitudinal plane. The PIR extends about one-third the way across the tooth and is straight. The bridge between the trigonid and talonid is wide. No crenulations are shown in Khomenko's illustration (Khomenko 1914, plate 1.20). Crenulations are absent or extremely gentle and an anteroexternal reentrant appears to be absent in the p4-m2s. The p3s of A. elongatus differ from the p3 of A. laskarewi in that A. elongatus has an AIR, a deeper PIR (and thus a narrower bridge between the trigonid and talonid), and crenulations in the reentrants. In addition, the p4-m2s of A. elongatus have crenulations on the anterior end of the talonid and may have anteroexternal reentrants. Comparisons with other species of Alilepus from North America. White (1991) recognized three species of Alilepus in North America: A. hibbardi, A. vagus Gazin, 1934, and A. wilsoni. He questionably assigned a fourth species, ?Alilepus browni (Hay 1921) to Alilepus. The temporal range of Alilepus hibbardi, A. vagus, and A. wilsoni is late Clarendonian (late Miocene) to late Blancan (late Pliocene). ?Alilepus browni is from the early Irvingtonian (early Pleistocene). Alilepus hibbardi White, 1991, is known from a complete skull and a total of five isolated teeth from Idaho, California, and Nevada. On p3, the PIR is generally shallow: it extends from about one-fourth to one-third the way across the tooth. On one tooth it is present as an enamel lake. The relative location of PIR is transverse or anterior to the PER. The PER extends about halfway across the tooth. There is a wide bridge between the trigonid and talonid. The AIR is lacking, although there is a slight indentation in the area of this reentrant on one tooth. The TH of the PER and the PIR may be smooth or lightly crenulated. Enamel of the AER is smooth. The p4-m2s of the holotype (IMNH 38695) have many light crenulations on the anterior end of the talonid and lack anteroexternal reentrants. In comparison, A. elongatus has a deep PIR located transverse to the PER, a distinct AIR, and an anteroposteriorly elongated trigonid. GSP 50959 is more heavily crenulated than specimens of A. hibbardi, but the degree of crenulations on GSP 34504 may be comparable. The p4-m2 of A. elongatus and A. hibbardi both have crenulated reentrants, but A. hibbardi lacks an anteroexternal reentrant, which is sometimes present on A. elongatus. Alilepus vagus Gazin, 1934, includes abundant remains from Nebraska, Idaho, and Washington (White 1991). Various authors have assigned the species to either Alilepus or Pratilepus. The three p3s illustrated by White (1991) have a PER extending at least halfway across the tooth and a PIR extending about one-third of the way across or forming an enamel lake. AIR is present or absent. White (1991) noted that dentally, A. vagus differed from A. hibbardi in the former having p3s with a deeper PER. Thus, differences between the p3 of A. elongatus and A. vagus are comparable to the differences between A. elongatus and A. hibbardi. Comparisons were not made between the p4-m2 of A. elongatus and A. vagus. Alilepus wilsoni White, 1991, is known from two fragmentary mandibles and 10 isolated p3s from Arizona. It is diagnosed as differing from other species of Alilepus in its smaller size and having a p3 with the PER with the inner portion widened and deflected anteriorly. This taxon has also been considered an immature Lepus or Hypolagus (White 1991). Presence of an elongated PER and lack of a PIR makes Alilepus wilsoni significantly different from A. elongatus. White (1991) questionably assigned Brachylagus browni Hay, 1921, (Hypolagus browni Dice, 1929) from Arizona to ?A. browni. The holotype of ?Alilepus browni is a fragmentary cranium, which "has no features that can be used for identification" (White 1991, p. 74). There are four isolated p3s of immature individuals. White's (1991) emended diagnosis of the species notes its small size and that the innermost part of the PER is not expanded and curved anteriorly as in A. wilsoni. Compared to the p3 of A. elongatus, the p3 of ?A. browni lacks a PIR or it may be present as an anteriorly located enamel lake, and AIR may be absent or present as a small indentation. Voorhies and Timperley (1997) assigned six isolated p3s from the Stewart Quarry, late Barstovian (ca. 13 Ma) of Nebraska, to Alilepus sp. The specimens are assigned to Alilepus, versus Hypolagus or Pronotolagus (both found in the same sediments) because they have either an enamel lake or a PIR. We agree with Voorhies and Timperley (1997) that these specimens conform to the generic diagnosis of Alilepus (White 1991), albeit an Alilepus with very primitive morphology (and certainly not comparable to the derived morphology of A. elongatus). Indeed, the Stewart Quarry specimens are quite similar in size and morphology to lithosympatric H. fontinalis. Voorhies and Timperley (1997) echoed White's (1991) proposal that Alilepus originated in North America, and later dispersed to the Old World, but added that the origin of the genus was at least 12 million years ago. Comparisons with Alilepus from Africa. A few p3s of Alilepus have been reported from Africa, but none have yet been described thoroughly. Cf. Alilepus was reported by Darwent (2007) from the late Miocene Lemudong'o Fauna, Kenya. Unfortunately, Darwent (2007) the material from Lemudong'o does not include any p3s. Haile-Selassie et al. (2004, figure 5) listed ?Alilepus in a composite faunal list from the late Miocene Adu-Asa and lower Sagantole formations, Middle Awash, Ethiopia. There was, however, no discussion of this taxon in the text, nor any illustrations. Wesselman et al. (2009) re-assigned the Adu-Asa material to Serengetilagus, and the Sagantole remains, including two p3s (dated at 4.85 Ma), to Alilepus sp. Photographs and brief descriptions were provided. The Ethiopian Alilepus appears distinct from the Siwalik and Kenyan specimens in (the Ethiopian specimens) possessing a very deep AR; PIR may be less deep than on the Siwalik/Kenyan Alilepus, but this is difficult to tell with certainty from the photographs. The length and width measurements of one Ethiopian p3 are given - 3.00 x 2.87 mm: the length is slightly shorter than the Siwalik and Kenyan p3s, but such small sample sizes preclude meaningful comparisons. The Ethiopian Alilepus needs to be more thoroughly illustrated and described before accurate comparisons can be made.
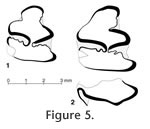
Comparable morphology suggests the two Kenyan Alilepus p3s illustrated in Figure 5 belong to the same species. The difference in size is similar to that between the two adult p3s of A. elongatus, and within the range of variation seen in other species of Alilepus. Morphology of the Kenyan specimens is comparable to that of A. elongatus, including relative width of the PIR, a lightly crenulated AER, and a lightly crenulated TN: neither Kenyan specimen has a crenulated TH. The major differences between the Kenyan and Pakistani specimens are that the Kenyan specimens have a less anteroposteriorly elongate trigonid, and the AIR may be absent. An incomplete p3 of a second specimen from the Lower Nawata Formation (KNM-LT 23179, Winkler 2003, figure 5.2B) also lacks an AIR and has a less elongate trigonid. Compared to the specimens illustrated in Figure 5, KNM-LT 23179 has a trigonid which narrows only slightly anteriorly, has a more anteriorly placed PIR, a wider bridge between the trigonid and talonid, and lacks crenulations in the reentrants. Differences between KNM-LT 22999 and KNM-LT 23179 are most likely due to morphological variation within a species, as both specimens are from the same general geographic area and are of comparable geologic age. Leporid p3 morphology is notably variable within a species, and the differences between A. elongatus and the Kenyan Alilepus may reflect morphological variability within a geographically widespread species. Because sample sizes are small, the Kenyan material is conservatively assigned as "near" A. elongatus. The Kenyan record is the likely result of dispersal of A. elongatus or a sister taxon from southern Asia to Africa during the late Miocene.
Pliopentalagus sp., Pliosiwalagus sp. or Caprolagus sp.
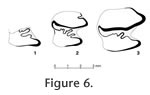
Occurrence. DP24, Pabbi Hills microsite, Soan Formation. Age. Ca. 1.7 Ma. Description. The crenulation pattern on the anterior border of the talonid of GSP 34829 (Figure 6.1), with two very distinctive crenulations, is strikingly different from the pattern seen on all other p4, m1, or m2s from the Potwar Plateau. There is a long anterior crenulation (extending about one-third of the width of the talonid), which has a small spur along its length. Distal to this crenulation is a second shorter one: it is about one-third the length of the first. In side view, the tooth curves posteriorly toward the base of the crown, suggesting it may be an m2 rather than m1, and not a p4. Talonid width of GSP 34829 is 2.40 mm. Discussion. GSP 34829 was compared with the talonids of p4 (for completeness), m1s, and m2s of other leporids having strong enamel crenulations. Among extant forms, this includes Pentalagus and Caprolagus. Among extinct taxa, it includes Pliopentalagus and Pliosiwalagus. Pentalagus furnessi, the Amami rabbit, is currently known only from the Amami Islands, southwestern Japan. The only fossil record of Pentalagus is two isolated teeth from the latest Pleistocene, Tokunoshima, Japan (Tomida and Otsuka 1993). Caprolagus hispidus, the hispid hare, lives along the southeastern foothills of the Himalayas in India, Nepal, and possibly Bangladesh (where it may be extinct). The only fossil record of Caprolagus was reported by Forsyth Major (1899) from the ?Upper Siwaliks of India. Patnaik (2002) re-assigned this material to the genus Pliosiwalagus Patnaik, 2001. Pentalagus has very large crenulations on the mesial borders of the talonids of p4, m1, and m2. Compared to the GSP 34829, the occlusal surface of Pentalagus is much more heavily and deeply crenulated with the deepest crenulations extending about 2/3 of the width of the talonid, and directed about 45 degrees to the anteroposterior axis of the tooth. Thus, we do not consider GSP 34829 to pertain to this genus. The extant genus Caprolagus may have well-crenulated anterior borders on the talonids of p4, m1, and m2. The degree and development of crenulations is variable, but may be close to that seen in the Pakistani specimen. However, as seen Figure 6.3, and based on our study of other specimens, the crenulations are not as pronounced on Caprolagus as on GSP 34829. Thus, although we cannot rule out the possibility that GSP 34829 belongs to Caprolagus, we consider it less likely than assignment to Pliopentalagus or Pliosiwalagus. Pliopentalagus includes material from Europe, China, and Afghanistan. Pliopentalagus dietrichi from Slovakia (Fejfar 1961; Daxner and Fejfar 1967) and P. moldaviensis from Moldavia (Gureev and Konkova in Gureev 1964) are both late Ruscinian in age. Chinese taxa include P. huainanensis (the oldest species of the genus, latest Miocene, MN 13; Jin 2004), P. dajushanensis (early Pliocene), and P. anhuiensis (late Pliocene) from Anhui Province (Tomida and Jin 2009; = spp. B and C, respectively, of Tomida and Jin 2002). Pliopentalagus huainanensis, P. dajushanensis, and P. anhuiensis represent a sympatrically evolving lineage showing gradual changes in several characters through time (Tomida and Jin 2009). The most recent species of Pliopentalagus is P. progressivus from Henan Province, dating to the latest Pliocene or earliest Pleistocene (Liu and Zheng 1997). Pliopentalagus nihewanensis was also described from China (Cai 1989), but Tomida and Jin (2005) have re-assigned this species to Trischizolagus. Pliopentalagus sp. was described from the early Pliocene Pul-e Charkhi locality, Afghanistan (Sen and Erbajeva 1995). This material, as well as specimens assigned to Serengetilagus by Sen and Erbajeva (1995), was originally allocated to Trischizolagus cf. maritsae by Sen (1983). Averianov and Tesakov (1997) returned Pliopentalagus from Pul-e Charkhi to Trischizolagus. GSP 34829 was compared with published descriptions and illustrations of all species of Pliopentalagus. The largest of the crenulations on GSP 34829 is significantly larger than any seen on the five illustrations of m1s (Daxner and Fejfar 1967) and 8 lower molariform teeth (Fejfar 1961) of P. dietrichi. No p4, m1, or m2s of P. moldaviensis Gureev and Konkova in Gureev, 1964, are available for comparison. Three mandibles of P. huainanensis were described and illustrated by Jin (2004, figures 2, 4). The p4-m2 of this species was diagnosed as having a distinct anteroexternal reentrant (PET in terminology of Jin 2004). Unfortunately, the trigonid is lacking on GSP 34829. The pattern of enamel crenulations on GSP 34829 is close to, but does not exactly match, the range of variation seen in P. huainanensis (e.g., Figure 6.2). The p4-m2 of P. dajushanensis and P. anhuiensis have basically the same enamel pattern on the mesial border of the talonid as P. huainanensis (Tomida and Jin 2009). Thus, the results of comparisons of GPS 34829 with P. dajushanensis and P. anhuiensis are the same as that with P. huainanensis. Pliopentalagus progressivus was described based only on one isolated p3 and one isolated p4 or m1 (Liu and Zheng 1997). It was distinguished by characters of the p3 and is the largest species of Pliopentalagus. It was considered the species of Pliopentalagus most similar to the extant taxon, Pentalagus furnessi. The p4 or m1 of Pliopentalagus progressivus was neither illustrated nor described, nor were its measurements given. Patnaik (2001, 2002) described two species of Pliosiwalagus based on small samples of Pliocene age (one locality about 3.5-4.5 Ma and the other about 2.5 Ma) from Himachal Pradesh, northern India. The genus is diagnosed by characters of p3, but is characterized also by possessing strongly crenulated thin enamel on p3-m2 (Patnaik 2002). The crenulation pattern seen on the thin enamel of GSP 34829 is within the range of variation of the enamel pattern on the p4-m2s of the illustrated specimens of Pliosiwalagus whitei (Patnaik 2001, figure 11a, b), however, the Pakistani specimen is too incomplete for definitive assignment to this genus.
Leporidae, genus and species undetermined
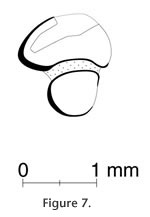
Occurrence. GSP 34510, 34511, 34828 are from DP8; GSP 39403, 34503, and 50954 are from Y457; GSP 50585 from Y581; GSP 49887, 49889, and 50269 are from Y908. Specimens from DP8 attributed to the local Tatrot Formation: all others are from the upper part of the Dhok Pathan Formation, Siwalik Group, Potwar Plateau, Pakistan. Age. Y457, 7.4 Ma; Y581, 7.15 Ma, and Y908, 7.0 Ma; and DP8, ca. 3.5 Ma. Discussion. Two small incomplete trigonids of p4, m1, or m2s (GSP 34510, 34828) are different in size and color and probably were not from the same individual. Both teeth lack an anteroexternal reentrant. However, the m3 (GSP 34511) is similar in size and color to GSP 34510, so these teeth possibly represent the same individual. The m3 (Figure 7) has a semi-circle shaped trigonid with the rounded portion anterior. The talonid is round and about half the size of the trigonid. The m3 measures 1.40 mm length, 1.24 mm trigonid width, and 0.66 mm talonid width. All of these dental remains (from locality DP8, ca. 3.5 Ma) are clearly Leporidae, but lack features diagnostic of Alilepus, and we hesitate to assign them at the generic level. Leporid post-cranial remains from the Hasnot area compare well with extant Leporidae of similar size and are likely referable to Alilepus. |
|