|
|
|
RESULTSLeporidae (Lepus)Published Reports and Identification Criteria. Early publications of the vertebrate fauna of the Anza-Borrego Desert listed Lepus and/or cf. Lepus without reference to particular specimens or localities (Downs and Woodard 1962; Downs and White 1968; Kurtén and Anderson 1980). In the first published study of the Anza-Borrego Desert fossil leporids, White (1984) listed no specimens definitively as 'Lepus', but identified three specimens as 'Leporinae, genus, and species indeterminate' because they could represent either Lepus or Sylvilagus. Subsequent statements attributed generic and tentative specific allocations to personal communications from John White. For example, Lundelius et al. (1987, p. 214) listed Lepus in the Anza-Borrego Desert at "about 2.0 Ma (White, personal commun., 1985)", and subsequently stated (page 220) "In the Vallecito Creek Local Fauna in the Anza-Borrego sequence . . . Lepus cf. callotis occurs at the base of the Olduvai subchron at about 1.9 Ma (White, personal commun., 1985)." In his major review paper on Miocene and Pliocene leporines, White (1991a:78) listed only two Blancan locality records for Lepus (Borchers in Kansas and Big Springs in Nebraska). A Lepus record at Anita, Arizona, indicated on White's (1991a:87) figure 14, was plotted in the transition zone between Blancan and Irvingtonian ages. The Anita fauna may be much older, is taxonomically and stratigraphically problematic, and needs full reexamination (Morgan and White 2005). White (1991a:87 figure 14) plotted two Lepus records as Irvingtonian (sensu Lundelius et al. 1987) in age, one from Inglis 1A in Florida, and another from the Anza-Borrego Desert, California. Our interpretation of the latest Lepus record in that figure is based on the caption statement, "The small horizontal lines indicate stratigraphic level in the Anza-Borrego Desert Section;" one such line appears at the top of the Lepus stratigraphic occurrence bar. However, the symbology is inconsistent elsewhere within the figure (e.g., the Aztlanolagus occurrence bar mistakenly includes a reference to the Anza-Borrego Desert Section). The 'ABD Lepus' mark is placed opposite the Coleman 2A (Florida) label and might also be interpreted as indicating the Lepus reported from there by Martin (1974). White did not discuss individual specimens in his (White 1991a) review. A total of four specimens from ABD were, at some time, referred in publication at least tentatively to Lepus. The documentary history, taxonomic, and stratigraphic complications of those specimens are discussed individually below. Taxonomic determination of isolated partial dentaries and teeth of North American leporines can be difficult or impossible. The enamel pattern and degree of enamel folding of the lower premolar (p3), the anteroposterior length of the tooth, and the i–p3 diastema length are the only characters suggested to permit differentiation of Nekrolagus progressus, some species of Lepus, and some species of Sylvilagus (e.g., Hibbard 1963; Dalquest 1979; White 1984, 1991a, 1991b; Dalquest et al. 1989). In the paedomorphic pattern of leporines, the posterior internal reentrant (PIR) is open lingually. Adult specimens of Nekrolagus, Lepus, and Sylvilagus typically have an enamel band along the lingual edge, closing the PIR. If the cement in the PIR is not continuous with that in the posterior external reentrant, a cement filled enamel lake or fossetid (Dalquest et al. 1989) is created, designated the 'Nekrolagus-pattern' (figure 1a); otherwise the PIR is continuous with the PER, resulting in the 'Lepus-pattern' (Hibbard 1963, figure 1b). Both patterns occur variously in Nekrolagus, Lepus, and Sylvilagus. The Nekrolagus-pattern was recorded in 88% (156 of 178 specimens) of N. progressus (Hibbard 1963), three of 22 specimens of S. webbi (White 1991b), one of 21 specimens of S. hibbardi (White 1984, 1991a), and one of four specimens of Lepus from Big Springs, Nebraska (White 1991a, figure 9). It is otherwise rare in Lepus and other Sylvilagus (Hibbard 1963). Specimen Descriptions and Provenience. ABDSP(IVCM) 113/V413 is a partial left dentary with p3 (Figure 1.1) and p4. Approximately 90% of the p3 occlusal surface is broken and missing although most of the enamel pattern in the cross-section of the broken area is visible. The anterior third of the tooth is broken below the alveolar rim, leaving the outline of the tooth enamel partially obscured, and the anterior reentrant ill defined. The anteroposterior length of the tooth is 3.7 mm. The i–p3 diastema length is 18.3 mm, although the anterior margin is incomplete. The bone appears to be diagenetically unaltered with no matrix attached to the bone and no visible permineralization or internal encrustation of gypsum or calcite crystals. Foramina and vacuities in the bone are partially filled with unconsolidated sand grains. The identification of specimen 113/V413 was given as "Lepus sp. cf. L. callotis" by Cassiliano (1994, p. 147). Specimen V413 shows none of the characteristics of preservation through protracted burial and permineralization as seen in specimens V1275 and V24889. Although this does not prove specimen V413 is not a fossil, it does leave open the possibility that it may be a modern specimen. Leporine bones are among the most abundant of recently deposited bones found on and in the surface sediments in ABD, and sometimes they are collected mistakenly as fossils. Once returned to the laboratory they can be difficult to separate from fossil specimens without detailed provenience information. Field notes are not known for most of the IVCM localities, so written and dated records of the circumstances of collection are not available for V413. ABDSP(IVCM) 113/V1275 is a partial right dentary with p3 (Figure 1.2), p4, m1, m2, and m3. The anteroposterior length of the p3 is 4.0 mm. The i–p3 diastema is missing. The specimen appears to be diagenetically altered and is encrusted with matrix. All openings are either permineralized or filled with cemented matrix. Field notes and written record of collection circumstances are unavailable for this specimen. Primary documentation (specimen catalogue cards) for ABDSP(IVCM) 113/V1275 includes two different field numbers, GJM 947 = (IVCM) Locality 27, and GJM 1024 = (IVCM) Locality 113; the two localities are physically separated by about 20 m. The date of collection, '26 April 1978', is consistent on all documents for this specimen, however, no other specimen in the current ABDSP database is listed with that date. All other specimens from locality 113 were collected on 24 November 1974. Locality 27 lists several dates of collection for the specimens, indicating separate survey and collecting events. The majority of the specimens from locality 27 are from disaggregated coprolites. Whether or not V1275 was recovered from a coprolite is unclear. Cassiliano (1994, p. 147) reported this specimen as 'Lepus callotis' under catalogue number 113/V4812 (=V1275, L.K. Murray specimen notes, 23 May 1996, on file at SRC). ABDSP(LACM) 1906/V24889 is a partial right dentary with p3 (Figure 1.3), p4, m1, and m2. The anteroposterior length of the p3 is 3.8 mm. The i–p3 diastema length is 17.3 mm, although the anterior bone margin is incomplete. The specimen appears diagenetically altered with matrix attached, permineralization, and crystalline growth present as on specimen V1275. The catalogue number of ABDSP(LACM) 1906/V24889 was misprinted in the caption of figure 6 (White 1984, p. 46) as "LACM 24839." The undated label in the specimen tray for this fossil has the words 'Lepus' and 'cf. L. callotis' printed on the taxon name line by two different hands with 'JAW' (J. White's initials, in his handwriting) on the same line. This fossil is the only ABD specimen with direct indication of identification to 'Lepus' by White. Although it is unclear to which identification the signature refers, either indicates that White at some time considered the specimen to be Lepus. ABDSP(IVCM) 110/V407 is a partial right dentary with p3, p4, and m1, and was reported as "Lepus sp." by Cassiliano (1994, p. 147). The specimen is no longer available for study. It was reinterpreted to be a modern specimen by the ABDSP curatorial staff, and according to the ABDSP database was removed from the collection and discarded. This removal occurred before the curatorial staff was aware that the specimen was included in Cassiliano's dissertation (1994). ABDSP documents indicate that specimen V407 was among the leporid specimens loaned to John White in 1979, although he did not include it in any of his publications. Subsequent publications by Cassiliano (1997, 1999) listed leporid specimens by taxonomic designation ('Lepus callotis', 'Lepus sp. cf. L. callotis', and 'Lepus sp.') but not by specimen number. Taxonomic Allocation. In the original description of the three fossil specimens under discussion, White (1984, p. 53) stated that the fossils were comparable to the mandibles of Lepus townsendii and Sylvilagus cunicularius and, "In enamel patterns of cheek teeth, specimens of S. cunicularius are indistinguishable from any North American species of Lepus or Sylvilagus. Thus if the only available characters to identify a specimen are size and enamel patterns on p3, the specimen cannot be referred with certainty to either Lepus or Sylvilagus." Using these criteria he concluded that the specimens were identifiable only to "Leporinae, genus, and species indeterminate." Subsequently, White (1991a:78) indicated that "larger size and the presence of folding of TN in PER [thin enamel on the posterior margin of the posterior external reentrant] could be used to diagnose S. cunicularius," and that mandibles could be identified as Lepus if the [i–p3] "diastema [is] 21 mm in length or longer," or if the p3 has the combination of "an anteroposterior length of 3.7 mm or larger" and enamel pattern similar to the predominant patterns of "L. californicus and L. townsendii, as shown in {f}igure 10." All three of the ABD fossil p3s have an anteroposterior length of 3.7 mm or greater. The two specimens with measurable (although incomplete) i–p3 diastemas each have a diastema length of less than 21.0 mm. Specimen V1275 has a cement-filled enamel lake (fossettid) separated from the PER by a dentine isthmus. Folding of thick enamel in PER (TH) of the three ABD p3s is 'simple'—"one small or shallow loop at the lingual margin of the central angle and/or a loop at the extreme lingual end of the re-entrant" to 'moderate'—"as in simple but with one or two additional small loops" (Dalquest et al. 1989, p. 299), while folding of TN is 'none' or 'simple'. Based solely on the criteria of White (1991a; enamel pattern on p3 similar to the predominant enamel patterns of L. townsendii and L. californicus as in his figure 10 and anteroposterior length of 3.7 mm or greater) the two specimens V413 and V24889 might be considered to be Lepus. However, V413 shares similar enamel characters (TH and TN in PER) with Nekrolagus progressus specimens reported by Hibbard (1963, p. 3, figures 1f, g, h, i), and falls within the size range of N. progressus (anteroposterior length of p3 = 2.7–3.8 mm, N = 38 specimens; White 1991a, table 3). Specimen V24889 shares similar enamel characters with Sylvilagus webbi. The p3 falls within the size range of S. webbi (anteroposterior length of p3 = 3.0–4.3 mm, N = 22 specimens; White 1991b, table 1); and its enamel pattern is similar to at least two S. webbi specimens (UF49841 and UF49847 in White 1991b figure 1). The enamel lake in specimen V1275 makes it a likely candidate for Nekrolagus (White 1991a) but an enamel lake is also found in some specimens of Lepus and Sylvilagus (Hibbard 1963; White 1991b). The incomplete diastema lengths of specimens V24889 and V413 approach 21 mm, one of White's (1991a) criteria separating Lepus from Sylvilagus. The two specimens also fall near the high end of the ranges of Nekrolagus progressus as well as S. webbi specimens from chronologically similar (Blancan) Florida sites; the diastema length range of Nekrolagus progressus is 16.8–18.5 mm (N = 3 specimens; White 1991a); and the diastema length range of S. webbi is 14.3–18.8 mm, (N = 7 specimens; White 1991b). All three ABD p3 specimens also appear to be similar in size and/or enamel pattern to the modern S. cunicularius specimen, UADZ5133 of White's (1984, p. 46) figure 6. Locality (IVCM) 113 is located stratigraphically near the base (1.95 Ma) of the Olduvai magnetic polarity chron C2n while locality (LACM) 1906 is above the top (1.77 Ma) of the Olduvai. The oldest age estimate that can be applied to locality (IVCM) 113 is between about 1.9 and 2.0 Ma, and for locality (LACM) 1906 between about 1.3 and 1.4 Ma. These two localities lie in the middle of the geographic and stratigraphic distribution of Sylvilagus hibbardi in the VCFC section, along with multiple sites listed on the database as containing elements diagnosable only to 'Sylvilagus or Lepus'. The estimated age of locality 113 reveals that specimens V413 and V1275 are those referred to by Lundelius et al. (1987, p. 214, 220; see above) as either "Lepus" or "Lepus cf. L. callotis." However, none of the ABD specimens has an enamel pattern remotely similar to the predominant enamel pattern for L. callotis as illustrated by White (1991a, figure 10), which shows a strong (sensu Dalquest et al. 1989) folding pattern of the TH enamel and moderate to strong folding of the TN enamel. The three fossil specimens discussed herein appear significantly larger than S. hibbardi, also reported from the associated strata, and may represent one or more different taxa. Based on criteria developed in previous discussions (e.g., Hibbard 1963; Dalquest 1979; White 1984, 1991a, 1991b; Dalquest et al. 1989), specimen V1275 appears similar to Nekrolagus, and specimens V413 and V24889 appear similar to Lepus and some large Sylvilagus. However, specimen V413 also appears to be a modern specimen while specimen V1275 suffers from imprecise provenience data. All three specimens lack sufficient diagnostic characters for incontestable designation as Nekrolagus, Lepus, or Sylvilagus. Well supported taxonomic identity and stratigraphic position are paramount for establishment of early records for these taxa. We therefore reaffirm White's original (1984) referral of these three specimens to 'Leporinae, genus and species indeterminate.' We searched the ABDSP fossil collection for other specimens possibly referable to Lepus, but found none with sufficient diagnostic characters for reference to that genus. ArvicolinaePublished Reports and Identification Criteria. Fossil specimens from ABD were referred previously to several arvicoline taxa including Ondatra idahoensis, Ophiomys parvus, Mictomys anzaensis, Lasiopodomys sp., Microtus (sometimes as M. californicus, sometimes with questionable allocation to that species, sometimes simply as 'Microtus sp.'), and Microtus (or Terricola) meadensis (Zakrzewski 1972; Repenning 1992; Repenning et al. 1995; Bell et al. 2004a). Only the Microtus and Lasiopodomys specimens are reviewed here. The recognized North American arvicoline fossil record consists predominantly of isolated teeth and partial tooth rows preserved in fragmentary (or, rarely, intact) lower jaws and maxillae. The lower first molar (m1) has long been recognized as the most useful tooth for determining taxonomic affinity in North American arvicolines. For some taxa the upper second and/or third molars (M2, M3) may also help to refine taxonomic allocation. Relatively few discrete morphological characters are preserved on isolated m1s, but systematically informative features include the presence or absence of roots, presence or absence of cementum in reentrant angles, the number of alternating triangles, differentiation in thickness of enamel on the anterior and posterior edges of triangles, and the relative closure of triangles with respect to each other and with other dentine fields on the tooth (posterior loop, anterior loop).
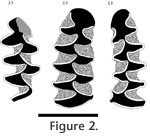
ABDSP(LACM) V24540 is an isolated, unrooted, right m1 (Figure 2.2); intact posterior loop with five closed triangles; sixth and seventh triangles are well-developed and confluent with each other and with the rounded anterior cap of the tooth. Enamel thickness is strongly differentiated on all triangles, with trailing edge enamel on each triangle thinner than that on the leading edge. Extensive cement is present in all reentrant angles. The anteroposterior length of the m1 is 3.4 mm. ABDSP(LACM) V24540 was originally published under locality '6814' (Zakrzewski 1972). Subsequent publications listed the specimen locality as '6683' (Repenning 1992) and '6686' (Repenning 1998). The primary data for this specimen are inconsistent. The specimen label, vial cork, gelatin capsule, and 1994 LACM database all have 'LACM 24540' and locality '6814' printed on them, as published originally by Zakrzewski (1972). The field notes, dated 22 January 1968, for locality 6814 (= Field Number HJG446) list "–fox lower jaw frag.–small pellet–rabbit skeleton?—Possible 2 rabbit skeletons or at least parts, as another cranium was close by & enclosed in jacket." The date of collection on the database catalogue for most specimens (including V24540) from locality 6814 is given as '13 April 1968', three months later than the date given in the field notes. When Charles Repenning began working on the ABD arvicolines in 1990, LACM curatorial staff discovered the date discrepancy for specimen V24540. In correspondence to Repenning LACM staff noted that the tooth was clean of attached matrix, implying it was a screenwashed specimen rather than an in situ discovery, and therefore probably was not from locality 6814. Multiple ABDSP(LACM) localities were screenwashed around 13 April 1968, including localities 1357 and 6683, and these were suggested to Repenning as possible sources of V24540. Locality 6683 is the site from which an edentulous jaw, V24649, referred to 'Microtus californicus?' by Zakrzewski (1972) was recovered (see below), so at least one other arvicoline specimen is known from that locality. Other localities (besides 6683 and 1357) also produced matrix for screenwashing around the same time and may be the source of V24540 (written correspondence between C. Repenning and LACM, 09 May 1990 to 22 June 1990). The locality and stratigraphic data for this specimen are seriously compromised. The specimen cannot, therefore, be considered in evaluations of the temporal range of arvicolines within ABDSP. The referral of specimen V24540 to locality '6686' (Repenning 1998, p. 52, footnote 9) is apparently a misprint of '6683'. Locality LACM 6686 is not an ABD locality. That locality number does not appear in any of the 1990 'arvicoline' correspondence between Repenning and LACM. ABDSP(LACM) 6683/V24649 is an edentulous left dentary. The specimen clearly represents an arvicoline rodent with ever-growing teeth (there are no root sockets). The approximate outline of the m1 is still discernible and indicates that there were likely five lingual and four labial reentrant angles. Locality and stratigraphic provenience data are not disputed. ABDSP(LACM) 68123/V24828 is an isolated, unrooted left m1 (Figure 2.3) with an intact posterior loop with three closed triangles; the fourth and fifth triangles are confluent with one another in a rhombus, but are closed from the anterior dentine fields. The sixth and seventh triangles also are well-developed, and broadly confluent with each other and with the rounded anterior cap. Cement is present in all reentrant angles. The trailing enamel is only slightly thinner than the enamel on the leading edge on each triangle. The anteroposterior length of the m1 is 3.55 mm. The locality for this specimen (locality 68123) is recorded in the 1994 LACM database, on field aerial photographs, and on a handwritten note (T. Downs, 29 August 1990) as being in the Borrego Badlands. The field notes (field number HJG487, recorded 06 February 1968) placed it in Beckman Wash, Borrego Badlands. Fossils from the site included "horse limb elements—rodent deposit—Neotoma." At least two sacks of matrix were collected on 08 February 1968 for screenwashing. Downs (handwritten note 29 August 1990) gave an estimated age for this locality as "0.5 Ma or later." An erroneous report that the specimen was from the "Vallecito Creek fauna" (Repenning 1992, p. 50) was apparently based on a written communication from J. White in 1990. We were not able to locate this correspondence, but all other sources of information regarding the position of this locality within ABD indicate it is in the top of the Borrego Badlands, approximately 80 km north of VCFC, in significantly younger sediments. There does not appear to be any major discrepancy in other primary data, therefore the specimen is still valid for biostratigraphic consideration, however, it was not recovered from within the VCFC. Taxonomic allocation. ABDSP(LACM) 1942/V8252 was originally identified as Microtus californicus? (Zakrzewski 1972), but was subsequently transferred to Lasiopodomys sp. (Repenning 1992). Specimens that conform to Repenning's morphological concept of Lasiopodomys in North America are significantly different in morphology of the m1 from the living representatives in Mongolia and Korea (see Repenning 1992 figure 11, and his discussion on pp. 46–48). The m1s of extant species of Lasiopodomys resemble those of many extant species of Microtus with five closed triangles. Recent molecular data also nest extant Lasiopodomys within crown-group Microtus (Galewski et al. 2006), supporting an argument for subsuming at least extant species of Lasiopodomys under the name Microtus. The extinct 'Lasiopodomys' deceitensis from North America is distinct (in morphology of m1) from other North American arvicolines, but was originally placed in the genus Microtus (Guthrie and Matthews 1971). Such placement may be justified, but the taxonomic affiliations of deceitensis are currently based only on molar morphology. Recent molecular analyses show at least weak support for a monophyletic clade of endemic extant North American Microtus (e.g., Conroy and Cook [2000] and Galewski et al. [2006]; but see Jaarola et al. [2004]). Based on ingroup commonality and outgroup assessment from those trees, the primitive morphology for m1 in North American Microtus was five closed triangles (unambiguous from the Conroy and Cook [2000] analysis; ambiguous in the Galewski et al. [2006] trees). In light of these data, the higher-level taxonomic affinity of 'Lasiopodomys' deceitensis remains uncertain (Bell et al. 2004a; Bell and Bever 2006). The morphology of ABDSP(LACM) 1942/V8252 is consistent with that of 'Lasiopodomys' deceitensis from various other localities in North America (reviewed by Repenning 1992) and was placed in taxonomic association with those specimens by Repenning (1992). Our illustration differs in detail from that provided by Repenning (1992, figure 11A), but we accept his identification of the specimen as a Lasiopodomys-like morphotype, and hesitatingly retain the use of 'Lasiopodomys' as a higher-order taxonomic name for specimens with that general morphology. In many North American faunas, this morphotype occurs in such low abundance that it likely represents atypical morphologies of other species (e.g., Microtus paroperarius; Bell et al. 2004a). Although Microtus paroperarius is not recorded from the Anza-Borrego Desert, the single specimen with a Lasiopodomys morphotype probably does not warrant formal taxonomic recognition at this time. The taxonomic assignments of ABDSP(LACM) V24540 have always implied close affinity with the extant California species, Microtus californicus. It was questionably referred to that species by Zakrzewski (1972). The questionable status was dropped by Repenning (1992) who provided a definitive referral to species. The question mark was returned by Repenning et al. (1995), and the specimen was listed as Microtus sp. cf. M. californicus by Repenning (1998). There are no fewer than 12 recognized extant North American Microtus species that share the general m1 morphology seen in extant M. californicus (Bell and Barnosky 2000). Without using modern biogeography as a tool to refine taxonomic allocation, there are no known morphologic features of this specimen that permit definitive species allocation. The broader complication of identification of isolated molars of Microtus species were discussed by Bell and Repenning (1999), Bell and Barnosky (2000), Bell et al. (2004a), and Bell and Bever (2006). The specimen is best referred to as a member of Microtus with five closed triangles on m1. ABDSP(LACM) 6683/V24649 is an edentulous dentary fragment. The taxonomic affinity of the specimen is poorly resolved. The original description (Zakrzewski 1972) identified it as 'Microtus californicus?' because the alveolus indicated an unrooted tooth, with reentrant and salient angles consistent with referral to Microtus. The argument was supported by the observation that the alveolus morphology was inconsistent with Mictomys, the only other arvicoline known at that time from ABD. The fact that the specimen could represent a different genus or species was noted in the original description (Zakrzewski 1972). The last three decades have seen an increase in the known diversity of arvicolines from ABD (Repenning 1992; Repenning et al. 1995), as well as western range extensions for taxa not previously thought to have occurred west of the Great Plains (e.g., Microtus paroperarius, Allophaiomys pliocaenicus; Gillette et al. 1999; Bell et al. 2004a, 2004b). These developments, combined with an increasing preference for reliance on diagnostic features of the fossils themselves, lead us to conclude that the most defensible identification for specimen V24649 is 'Arvicolinae.' ABDSP(LACM) 68123/V24828 was identified as Terricola meadensis by Repenning (1992). The morphology of the specimen is certainly consistent with the type population of 'Pitymys' meadensis from the Cudahy fauna in Kansas (Hibbard 1944; Paulson 1961), and we agree that the species assignation is correct. Higher-level taxonomy for meadensis has been a source of confusion and contention for decades. Recognition of distinct 'pitymyine' species groups (variously placed in the genera or subgenera Pitymys, Pedomys, Terricola) was commonplace until the emergence of molecular data that universally found these groups to be polyphyletic (e.g., Moore and Janecek 1990; Conroy and Cook 2000). Based on all available data, the specimen is best considered Microtus meadensis (see discussion in Bell et al. 2004a). |
|