|
|
|
SYSTEMATIC PALEONTOLOGY
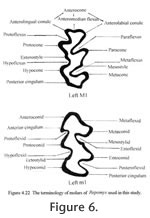
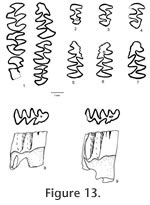
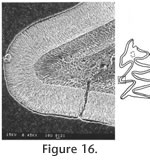
Order RODENTIA Bowdich, 1821 The terminology for the cusps and lophs follows Tomida (1987, figure16) except that the stylar cuspule associated with the anteroloph is named as the parastyle for upper molars, and the stylar cuspule associated with the anterolophid in the lower molars is named as the metastylid.
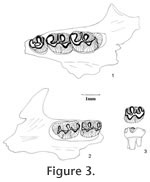
Peromyscus hagermanensis
Hibbard, 1962
The M1 has five distinct cusps: anterocone, protocone, hypocone, paracone, and metacone. The protocone and hypocone are located slightly anterior to the position of the paracone and metacone, respectively. The anterocone is broad, weakly bilobed, asymmetrical with a prominent labial cusp and a variably developed (small to indistinct) lingual conule. A shallow anteromedian flexus is distinct between the labial and lingual sides of the anterocone in unworn or slightly worn teeth; the flexus gets shallower with advanced wear and disappears with late wear. One specimen (UALP 22916 from Locality 9504) has a unicusped anterocone. The protocone has a long anterior arm directed toward and connecting with the anterocone and a short posterior arm directed obliquely to join the mure. The short anterior arm of the hypocone is directed toward the paracone and connects with a mesoloph and the mure. It is not in linear alignment with the paraloph. The posterior arm of hypocone joins the metaloph and continues labially as the posterior cingulum, and the posterior cingulum is distinct in unworn and moderately worn specimens but becomes indistinct in well-worn ones. The paracone and metacone are conical. Wear on lingual cusps is medial, and wear on labial cusps is posteromedial. A minute parastyle is present on 70% of M1s (Table 1), joining the paraflexus. A short anteroloph connecting the anterocone with the parastyle is present in 5% of the specimens. Sixty-three percent of M1s have a minute mesostyle, which may or may not connect with a mesoloph. Sixty-five percent of M1s present a mesoloph. One specimen from Locality 8197 (UALP 21251) has a minute protostyle. No enterostyle or enteroloph is seen on any of the M1s. Table 1 lists the frequencies of styles and lophs of M1s from each locality. Each M1 has three distinct roots (anterior, lingual, and posterior). None of the M1s developed accessory rootlets. The M2 is smaller than M1, with four prominent cusps: protocone, hypocone, paracone, and metacone. The anterocone is lost. Instead, a broad anterior cingulum is present, joining with the long anterior arm of the protocone near or slightly lingual to the midline. The labial half of the anterior cingulum is separated from the paracone until it reaches the labial margin. The short posterior arm of the protocone joins the paraloph and central mure. The paraloph and anterior arm of the hypocone are not in alignment. A posterior arm of the hypocone is present in unworn and moderately worn specimens. The development of styles and lophs are similar to those in M1 except that four out of five M2s from Locality 9602 and two out of nine from Locality 9504 have both anterior and posterior paralophs (Figure 3.3; paralophule I and II in Lindsay 1972, figure 40). This structure is not observed in any of the M1s. Each M2 has three roots. The M3 is well reduced relative to M1 and M2. A narrow anterior cingulum is present. The protocone and paracone are prominent, whereas the hypocone and metacone are minute or indistinct. A thin posterior cingulum is distinct on slightly worn specimens. The paracone has an anterior paraloph connecting with the anterior cingulum and a posterior paraloph that joins the posterior arm of the protocone to form a large enamel islet between the paracone and protocone. This anterior paraloph is present in all M3s. The M3s have two or three roots, with the medial and posterolabial roots fused in the teeth with two roots. The m1 is narrow anteriorly in occlusal view. The anteroconid is broad, weakly bilobed, with the lingual conule slightly larger than the labial conule. A short anterior cingulum almost closes the protoflexid. Seven of 41 m1s (17%) have a unicuspid anteroconid, resulting in an oblique occlusal outline. The principal cusps alternate in position, with the metaconid and entoconid placed anterior to the protoconid and hypoconid, respectively. The anterior arm of the protoconid is directed anteriorly and connected to the metalophid and a short posteriorly directed anterolophid from the anteroconid slightly labial to the median plane. The long posterior arm of the protoconid is directed toward and aligned with the entolophid regardless of the wear. The metalophid and entolophid are both short and straight. The anterior arm of the hypoconid is relatively short, joining the entolophid near the midline. The posterior arm of the hypoconid is longer, directed toward the posterior midline where it joins the posterior cingulum. The posterior cingulum is prominent and relatively heavy, oriented transversely. The mesostylid is present on 14% of m1s, and a mesolophid is present on 11% of the specimens (Table 1); both are less frequent than in M1s. No metastylid is seen on the m1s. Only one specimen (22956 from Locality 9601) shows an ectostylid and a long ectolophid. Each m1 has two roots without any accessory rootlets. The m2 has a rectangular occlusal outline. The cusps and lophids are similar to those in m1 except that the anteroconid is absent. There is a shallow but distinctive posterolabial sulcus between the hypoconid and the prominent posterior cingulum, as in m1. A mesostylid is present on 17% of the m2s (seven out of 42), and different from the other localities, Locality 9602 has 45% of m2s (five out of 11) with the mesostylid. No mesolophid is seen on any m2. Only one m2 (UALP 23010 from Locality 9602) has a distinct ectostylid, but it has no ectolophid. Each m2 has two roots. The m3 has a prominent protoconid and metaconid, and a smaller hypoconid. The entoconid is indistinct or absent. An anterior cingulum is developed on the labial side of the tooth, anterior to the protoconid. The posterior cingulum is usually reduced, forming a thin enamel wall that joins the hypoconid and the metaconid on the lingual side of the tooth. Each m3 has two roots. Discussion. It was noticed that a few m1s have an oblique occlusal outline and differ from the majority of m1s in this fauna. These specimens are UALP 21262 and 21265 from Locality 8197, UALP 22921 from Locality 9616, UALP 22903 from Locality 9504, UALP 22993 and 23001 from Locality 9602, and UALP 22956 from Locality 9601. Each of these teeth is narrow anteriorly with a unicuspid anteroconid. Their averaged anteroposterior length is 1.57mm, and transverse width is 0.98mm. These measurements are very close to the other m1s of P. hagermanensis from the fauna. They are also close to the 14 specimens of Peromyscus hagermanensis from the Duncan and 111 Ranch local faunas of Arizona, which have an average length of 1.58 mm and width of 0.99mm (Tomida 1987, table 20). One-third of the m1s from the 111 Ranch l.f. also have a unicuspid anteroconid. Therefore, the seven specimens from the Panaca l.f. mentioned above are here assigned to P. hagermanensis. Reithrodontomys, another brachydont cricetid, often has a unicuspid anteroconid on m1; however, members of this genus are distinctly smaller in size. Lindsay (1972) found that the differential character of living Peromyscus from Copemys is the alignment of the paraloph-anterior arm of hypocone in the M1-M2 and entolophid-posterior arm of protoconid in the m1-m2 in Peromyscus. In the M1s and M2s of P. hagermanensis from the Panaca fauna, the paraloph and anterior arm of hypocone are apparently offset. However, this alignment does occur in the m1s and m2s. After checking the upper molars of P. hagermanensis from the type locality (UMMP 52739, Zakrzewski 1969) and from the Duncan and 111 Ranch faunas in Arizona (Tomida 1987), the author found that the nonalignment in upper molars is quite common in those samples. The modern species Peromyscus eremicus also has non-alignment of the lophs (Lindsay, personal commun., 1999). Therefore, the only character that consistently differentiates Peromyscus from Copemys is the alignment of entolophid-posterior arm of protoconid in the lower molars of the former. The alignment in lower molars and nonalignment in upper molars are also seen in two species of Copemys: C. valensis (Shotwell 1967) and C. vasquezi (Jacobs 1977). Korth (1994) considered them ?Copemys as they could be justifiably included in Peromyscus. As Tomida (1987) suggested, P. hagermanensis is closer to Copemys in this feature than the other species of Peromyscus. Brachydont cricetids have very similar dental morphology. Tomida (1987) gave some characters to distinguish Onychomys, Peromyscus, Calomys (Bensonomys), Reithrodontomys, and Baiomys. Peromyscus hagermanensis from the Panaca fauna can be differentiated from Onychomys by its smaller size and narrower reentrant valleys; from Calomys (Bensonomys) by its weakly bilobed anterocone, shallower anteromedian flexus, higher and steeper cusps, and lack of accessory rootlets; from Reithrodontomys by its larger size, and from Baiomys by its larger size, wider lower incisor, and weakly bilobed anterocone. The Hemphillian Peromyscus antiquus has a significantly larger size compared to P. hagermanensis. P. kansasensis (Hibbard 1941) from the Blancan Rexroad fauna has a similar size, but it lacks accessory styles or lophs. P. sawrockensis (Hibbard 1964) from the Sawrock Canyon l.f. is larger and lacks accessory styles or lophs. P. baumgartneri from the Rexroad l.f. (Hibbard 1954) and P. cragini from the Cudahy l.f. (Hibbard 1944) are smaller and either lack styles or retain minute vestiges of them. P. nosher from the early Blancan White Bluffs (Gustafson 1978) l.f. has a distinctly bilobed anteroconid on m1, and a broad and smooth connection between the posterior cingulum and the hypoconid of m2. Dalquest (1978) erected a new species, P. beckensis, for the brachydont cricetids from the early Blancan Beck Ranch l.f. and gave the diagnosis of this species as "having m1 slender, narrowed anteriorly, m3 unreduced, mental foramen dorsal, and dentition brachydont." Only the dorsally located mental foramen can distinguish P. beckensis from the other species of Peromyscus. The M3s from the Panaca fauna are slightly larger than those from the Duncan and 111 Ranch faunas. No M3 of P. hagermanensis has been found at its type locality. It should be noted that there are some slight differences in Peromyscus hagermanensis among the Panaca localities. Four M1s from Locality 9702 have no styles or lophs. Three of them are well worn, and the less worn specimen shows a distinctively bilobed anterocone with a shallow but very wide anteromedian flexus. This character is not seen in the other M1s. Four out of five M2s (18%) from Locality 9602 have two paralophs, whereas the majority of M2s from the other localities only has a posterior paraloph. Since the other molars from these two localities are very similar to those of P. hagermanensis, I consider these abnormalities of M1s from Locality 9702 and M2s from Locality 9602 to be intraspecific variations. Occurrence. Rodent Ravine section: 9702, 9616, 9619; Limestone Corner section: 8197; Brown Bone Beds: 9620, 9621; Rodent Hill section: 9504, 9601, 9602, in the Fm., southeast Nevada; Duncan and 111 Ranch local faunas, Gila Conglomerate, southern Arizona; Hagerman l.f., Glenns Ferry Formation, Idaho. Range. Blancan.
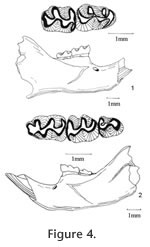
Genus ONYCHOMYS Baird, 1858
M Description. The maxillary from Locality 9504 is moderately worn. Posterior margin of the incisive foramina extends to approximately the anterior margin of M1. The anterocone is asymmetrical with a prominent labial conule. Presence of a lingual conule cannot be determined due to the wear. Labial cusps are located across from the posterior side of lingual cusps. Three molars do not have any styles or lophs. Both M1 and M2 are distinctively larger than those of Peromyscus hagermanensis (Table 3) and have wider reentrant valleys; however, the M3 is the same size as that of P. hagermanensis. M2 and M3 have a long labial anterior cingulum. The isolated M1 from Locality 8197 is broken at the posterior cingulum, but its overall size is comparable to the M1 of the maxillary from Locality 9504. This tooth is unworn, with a broad, slightly asymmetrically bilobed anterocone. The lingual conule is slightly lower than the labial conule. The cusps are high and steep. It has a minute parastyle. There is no mesoloph. The reentrant valleys are wide. The mure is longer and higher than that of P. hagermanensis. The two isolated m1s from Locality 9504 and 9601 are larger and more robust than most m1s of P. hagermanensis (Table 3). The anteroconid is short, narrow, symmetrical, and unicuspid. The anteroconid is slightly smaller than the more prominent alternating cusps. The protoflexid is slightly closed due to a low anterior cingulum. The posterior cingulum is relatively short, terminating before reaching the posterior lingual corner of the tooth. Both specimens have wide reentrant valleys, high and steep cusps, and high mures. They do not have any stylids or lophids. The m2 from Locality 9620 is larger than the m2s of P. hagermanensis. The cusps are alternating but not as strongly as in P. hagermanensis; the mure is relatively long, and the reentrant valleys are wide. The short posterior cingulum resembles a small lingual posteroconid. It has no stylids or lophids. Discussion. These specimens are referred to the genus Onychomys because of their larger sizes, steeper cusps, near absence of accessory styles and lophs, and wider reentrant valleys compared to Peromyscus. There are two recent grasshopper mouse groups in North America, O. leucogaster and O. torridus, one larger than the other. The earliest known species of Onychomys is O. martini from the late Hemphillian Edson Quarry of Kansas (Hibbard 1937). The early Blancan species are O. gidleyi Hibbard 1941 and O. larrabeei Hibbard 1953. Hibbard (1953) named O. larrabeei from the Sawrock Canyon l.f. of Kansas based on the sizes of molars; however, Carleton and Eshelman (1979) found that the size of O. larrabeei fell within the range of O. gidleyi, and therefore, they considered O. larrabeei as the junior synonym of O. gidleyi. The diagnostic characters on the species of Onychomys are focused on the lower jaws, especially the reduction of m3 and the development of capsular process on the dentary. Compared to the type specimen of O. martini (KU 3850, a right maxillary with M2-M3), UALP 22895 is very similar to it in occlusal view and size. The two m1s from the Panaca fauna are smaller than those of O. gidleyi or O. larrabeei but well within the range of smaller species O. hollisteri Carleton and Eshelman 1979 and O. bensoni Gidley 1922. On the other hand, the m2 from Locality 9620 is close to the m2s of O. gidleyi in size (Table 3). Carleton and Eshelman (1979) considered that O. gidleyi and O. pedroensis Gidley 1922 were in the same lineage giving rise to the recent species O. leucogaster. The size of the maxillary from Locality 9504 is very similar to those of O. pedroensis from the 111 Ranch l.f. of southern Arizona (Tomida 1987). Therefore, there could have been two groups of Onychomys living in Panaca at the early Blancan. The larger specimens may represent a species in the group of O. martini, O. gidleyi, and O. pedroensis, whereas the smaller one could be close to O. hollisteri or O. bensoni. Occurrence. Rodent Hill section: 9504, 9601, Limestone Corner section: 8197, and Brown Bone Beds: 9620, in the Fm., southeast Nevada. Range. Early Blancan to Recent.
Tribe Neomyini Merriam, 1894 Emended diagnosis. Neomyine with slender, high-crowned but rooted cheek teeth, having relatively thick enamel. There is no enamel islet on M1 or M2. M3 has two labial reentrants in an "E" shape when unworn or slightly worn, a single anterolabial reentrant in an "F" shape when moderately worn, or no reentrant but an anterior enamel islet when worn. Posterior enamel islet present or absent on M3. Keyhole-shaped m3 with an anterolingual reentrant and a short anterolabial cingulum that are lost with wear; anterior enamel islet present or absent on m3.
In the following description, the terminology of cheek teeth follows Tomida (1987) as shown in Figure 6. Since the occlusal view changes considerably at different level of wear, the degrees of wear are used for description as follows: unworn—no wear and roots not yet developed; slightly worn—wear is less than one-third of original crown, and roots are starting to develop but very short; moderately worn—crown is about one-third to half of original height, and roots are open; worn—crown is lower than half of original height, and roots are closed.
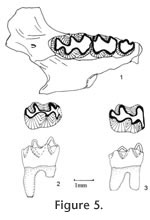
Repomys panacaensis
May, 1981
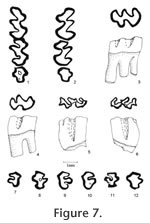
Material examined. UALP Locality 8197: M2s: 21182-21184; M3s: 21185-21188; 21190, 21191; right mandible with m1-m2: 21160; m1s: 21201-21203; m2s: 21205, 21211, 21212; m3s: 21213, 21217, 21425. Locality 9502: M1: 21400. Locality 9504: partial skulls with two incisors and two complete dentitions: 21658; SBDM 1; a partial skull with two incisors and left M1-M3: 21657; maxillaries with M1-M3: 21372, 21724; maxillaries with M1-M2: 21371, 21373, 22510, 21659-21663; maxillary with M2 and alveolus of M1: 21726; M1s: 21666-21668, 21670, 21727-21730; M2s: 21671-21673, 21733-21739, 21386; M3s: 21388-21391, 21741-21745, 21675-21684; mandibles with m1-m3: 21370, 22509, 21700, 21701; mandibles with m1-m2: 21702-21707, 21725, 21746; mandible with m1 and m3: 21708; mandibles with m3 and alveoli of m1 and m2: 21710, 21711; m1s: 21685-21691, 21747, 21748, 21376-21379; m2s: 21692-21694, 21749-21755, 21380-21382, 21384; m3s: 21756-21759, 21695, 21697, 21761, 21393-21395. Description. UALP 21657 preserved two upper incisors in the partial skull. The incisors are strongly curved, and the anterior parts point downward, almost perpendicular to the premaxillary. The incisors are smooth and slender, with a width of 1.15 mm. The diastema between the incisor and M1 is 7.80 mm. Measured from SBDM 1, the width of two nasal bones at the widest points is 3.15 mm, and choana is 2.45 mm wide. The posterior border of the incisive foramina terminates at the anterior or medial part of M1 at a point slightly more posterior than the posterior border of the incisive foramen of Neotoma. The anterior palatine foramen is 6.85 mm long, and it ends between the anterior and lingual roots of M1. The posterior palatine foramen is at the position of the posterior root of M2. The averaged alveolar length of M1-M3 from eight measurements is 5.33 mm. M1 is very high-crowned, as tall as it is long in slightly worn teeth. It has five major cusps: anterocone, protocone, paracone, hypocone and metacone. The anterocone is asymmetrically bilobed in unworn or slightly worn specimens, with the labial conule slightly larger than the lingual. The anteromedian flexus is very shallow, and it is lost at the early stage of wear (less than one-third of the original height of crown). The anterostyle on the posterior arm of the anterocone slightly constricts the paraflexus, and the mesostyle on the posterior arm of the paracone slightly constricts the metaflexus. These two small styles are obliterated at the moderately worn stage, and they never join the opposing enamel to form an islet prior to the loss of metaflexus through wear. In slightly worn specimens, the protocone is located slightly anterior to the paracone, and the hypocone is slightly anterior to the metacone. In well-worn specimens, the hypocone and metacone are nearly transversely opposed. The protocone is differentiated from the anterocone by a shallow protoflexus, and this protostria persists to just above the base of the crown. Posterior arm of the protocone may be separated from the anterior arm of the paracone in unworn specimens, but they become confluent with moderate wear. A small enterostyle is present on the posterior arm of the protocone in unworn to slightly worn specimens. A broad and deep hypoflexus separates the protocone and hypocone. Anterior arm of the hypocone is confluent with the paracone, and its posterior arm is confluent with the metacone. The paracone is separated from the anterocone by a deep, posterolingually directed paraflexus. It is separated from the metacone by a deep metaflexus. The metaflexus is directed posterolingually on unworn and moderately worn specimens and changes to a more transverse direction with wear. Both parastria and metastria continue to the base of the crown. All of M1s have three well-developed roots: anterior, lingual, and posterior. They are similar in size. M2 is similar to the M1 except for its shorter length and root condition. The shortness results from the loss of anterocone. On unworn M2s, the anterior cingulum is asymmetrically bilobed (Figure 7.5-7.6), but there is no structure homologous to the anterostyle of M1, even on young individuals. The mesostyle between the paracone and metacone is as distinct as in M1. The majority of M2s have three roots (Table 6): two anterior and one posterior. The anterolingual root is the strongest, and the anterolabial root is reduced. A few M2s (9%) with two roots have the two anterior ones fused, forming an L-shaped root. M3 is well reduced, and the occlusal outline changes dramatically at different wear stages. In unworn specimens, two labial reentrants are present so that the occlusal surface is E-shaped (Figure 7.9). The posterior reentrant is usually shallower than the anterior reentrant. Anterior cingulum is developed. The protocone is distinct, and the paracone is large. The hypocone is smaller than the protocone but is usually distinct until moderately worn; it is separated from the protocone by a shallow hypoflexus. The metacone is indistinct or absent. In slightly worn specimens, the hypocone and posterior cingulum are reduced, forming a rounded posterior heel. With wear, the posterior cingulum connects to the paracone closing the metaflexus to form a smaller and shallow posterior enamel islet (Figure 7.10-7.11). The enamel wall of this islet is very thin, unlike that of R. maxumi. Also with wear, the paracone may connect to the well-developed anterior cingulum to form a large and shallow anterior enamel islet that may persist until moderate wear (Figure 7.8). Alternatively (in 14% of the M3s, nine out of 65), the opposing enamel walls of the paraflexus never connect to form an islet, and the occlusal view results in an "F" shape (Figure 7.7). With greater wear the anterior and posterior islets are lost (Figure 7.12). The root condition of M3 is highly variable: most M3s have two roots (68%), some have only one root (24%), and a few retain three roots (8%; Table 6). The bases are fused on some of the M3s with two roots. The M3s with one root have a deep groove on either the labial or lingual side. The lower incisor is located labial to the m2 with the incisor cap placed between the coronoid process and the condyle on the labial side of the ascending ramus. The capsular process is prominent, forming a shelf that lacks a distinct knob. The masseteric crest extends anteriorly to a position beneath the anterior side of the protoconid of m1. There is a well-developed but short shelf lacking a knob at the anterior end of the masseteric crest. The mental foramen is relatively high on the dorsal-labial side of the dentary, well separated from the masseteric crest and the anterior root of m1. This position is similar to that of Neotoma but lower than that of Sigmodon, where the mental foramen is higher and located on the dorsal surface of dentary. The coronoid process of R. panacaensis curves posteriorly, and the condyle faces upward. The distance between the coronoid process and the condyle is 3.35 mm on a single measurable specimen (UALP 21701). The other measurements of these mandibles are included in Table 5. The m1 is hypsodont, as tall as long in unworn specimens. It has five cusps: anteroconid, protoconid, metaconid, entoconid, and hypoconid. The anteroconid is single-cusped with an anterolabial cingulum directed posterolabially. The anterior cingulum is lost after moderate wear. The metaconid is distinct but smaller than in R. gustelyi and R. maxumi. It confluently connects to the anteroconid and protoconid. The entoflexid is shallow; its apex projects anteriorly in unworn specimens, anterolabially in slightly worn specimens, and transversely in well-worn specimens. Anterior arm of the protoconid connects to the anteroconid, and the posterior arm is confluent with the anterior arm of the entoconid even in unworn specimens. The anterior arm of the entoconid joins a small mesostylid in unworn and slightly worn specimens, slightly constricting the apex of the entoflexid. The hypoconid and protoconid are labially separated by a broad and moderately deep hypoflexid, with the apex directed anterolingually. The anterior arm of hypoconid confluently joins the labial side of entoconid. A small ectostylid, smaller than mesostylid, projects anterolabially on the anterior arm of hypoconid in unworn and slightly worn specimens, similar to R. arizonensis. The protostriid is very shallow, and the hypostriid is deep but well above the base of the crown. The entostriid is slightly deeper than the posterostriid. The lingually directed posterior cingulum is very well developed; it is broadly confluent with the hypoconid. The posteroflexid is slightly shallower and much narrower than the entoflexid. The m1 has two distinct roots: anterior and posterior. Two specimens (UALP 21687 from Locality 9504 and 22718 from Locality 9602) have a tiny rootlet between the anterior and posterior roots. The m2 is similar to m1 except it is shorter. The anterior cingulum is relatively broad, with a short labial extension anterior to the protoconid. The entoflexid is directed anterolabially in all specimens. The posterior cingulum is longer in unworn and slightly worn specimens than in well-worn specimens. The enamel wall on the anterior side of posterior cingulum is very thin in unworn and slightly worn specimens, like that of R. arizonensis. The protostriid is very shallow; the hypostriid and entostriid are very deep but terminate above the base of the crown; the posterostriid is very shallow. The m2 has two distinct roots as in m1. The m3 is reduced relative to m1 and m2. It is keyhole shaped in occlusal view in moderately worn specimens. In unworn specimens, a shallow entoflexid is present, with the apex pointing anterolingually. An anterolabial cingulum is present in slightly worn specimens and absent in moderately worn specimens. The posterior heel represents the greatly reduced hypoconid; it is deflected lingually. The posterolabial reentrant (hypoflexid) is very broad and shallow, as is the posteroflexid. The entostriid is very shallow; the posterostriid and hypostriid are deeper, with the hypostriid nearly reaching the base of the crown. More than half of m3s in this sample have two roots as in m1 or m2 (66%), some partially fused, but the others have only one root, with the anterior and posterior ones fused medially or labially. Discussion. Three other species of Repomys have been reported in North America: R. gustelyi May 1981 from the late Hemphillian Warren l.f. of California, R. maxumi May 1981 from the early Blancan Maxum l.f. of California, and R. arizonensis Tomida 1987 from the late Blancan of 111 Ranch l.f. of Arizona. As May (1981) stated, R. panacaensis can be differentiated from R. gustelyi or R. maxumi by its smaller size. The M1 and M2 are very similar among these species; however, the M3s of R. panacaensis frequently present an anterior enamel islet, which is not seen in the other two species. The M3 of the latter two species is characterized by a deep anterior reentrant, but this reentrant never constricts to form an islet. Also, the posterior reentrant is deeper in the M3 of R. panacaensis compared to those of R. gustelyi or R. maxumi at the same stage of wear. In unworn or slightly worn M3s of R. panacaensis, this posterior reentrant forms the shape of an E in the occlusal view instead of an F that is seen in the other two species. Comparison of root condition of M3 between these three species is difficult because it is highly variable in R. panacaensis and only one specimen of R. gustelyi and four specimens of R. maxumi are available. Larger samples are needed to conclude if there is a trend in the root reduction on M3 from R. gustelyi and R. maxumi to R. panacaensis. The apex of entoflexid in m1 is another important character to distinguish R. panacaensis from R. gustelyi or R. maxumi (May 1981). In R. panacaensis, this reentrant only points anteriorly in unworn specimens and gradually changes its direction transversely in advanced wear. However, in the latter two species, the entoflexid is deep, and the apex is directed strongly anteriorly even in worn specimens. This configuration makes the metaconid rather separated from the anteroconid in the latter two species. The anterolabial cingulum in m1 is also obliterated with moderate wear in R. panacaensis, whereas it continues in more advanced wear in the other two species. The m2 and m3 are similar in the three species except for their sizes. The anterior enamel islet on m3 is preserved until moderate wear in R. panacaensis, similar to that in R. gustelyi and R. maxumi. When Tomida (1987) erected the species R. arizonensis, neither an M3 nor an m3 was found. He listed the following characters of R. arizonensis to differentiate it from R. panacaensis: 1) size is significantly larger; 2) the lingual reentrant on m1 extends further across the tooth, and the metaconid is not as close to the anteroconid; 3) posterior cingulum of m1 is better developed; 4) M2 has only two roots, with anterior and lingual roots fused, forming an L-shaped large root. It is noticed that when this comparison was made, only a small sample and moderately well worn specimens of R. panacaensis were available (see May 1981). With more specimens and unworn teeth available, there is only one character of those listed above that can distinguish R. arizonensis from R. panacaensis: the metaconid on m1 of R. arizonensis is better developed and less close to the anteroconid. The sizes of these two species are not significantly different, and on unworn or slightly worn m1s, the posterior cingulum in R. panacaensis is as developed as in R. arizonensis. The majority of M2s of R. panacaensis have three roots, but some of them are also reduced to two roots, and the anterior one is L-shaped. Since only one M2 of R. arizonensis was available for root identification (Tomida 1987), a larger sample is needed to make this character secure for differentiation. In addition to the metaconid on m1, two other characters may be added to distinguish these two morphologically similar species: 1) the metaflexid on m1 is deeper in R. panacaensis than in R. arizonensis at the same stage of wear. This configuration makes the metaconid more distinct from the anteroconid in R. panacaensis; 2) the hypoflexid on m1 is broader and shallower in R. panacaensis than in R. arizonensis at the same stage of wear. The apex of this flexid in R. panacaensis points more transversely. This character is seen in more primitive species like R. gustelyi. R. panacaensis could give rise to R. arizonensis, but the discovery of M3 and m3 of R. arizonensis will be very important and useful to understand the phylogenetic relationship between these two species. Repomys can by readily distinguished from two other similar hypsodont cricetines, Galushamys and Pliotomodon, by the absence of enamel islet on M1 or M2. Pliotomodon, as suggested by Jacobs (1977), is probably an immigrant close to European Ruscinomys hellenicus. Repomys differs from Neotoma or Neotomodon by its F-shaped M3 and keyhole-shaped m3. Based on the record of Peromyscus cf. P. pliocenicus from the early Hemphillian of Oregon, which starts to develop hypsodonty and has a single cusped anteroconid on m1, May (1981) suggested that Repomys was likely derived from P. pliocenicus. The M3 of P. pliocenicus has an E-shaped occlusal view, and the m3 is S-shaped with a deep posterior lingual reentrant (May 1981, figures 2c, 4). Both M3 and m3 of Peromyscus cf. P. pliocenicus have a primitive pattern compared to Repomys, but they are not quite reduced. Korth (1994) supported this P. pliocenicus-Repomys hypothesis, but he suggested that P. pliocenicus should be referred to a genus other than Peromyscus because its crown height is greater than any other known species of this genus. It is noticed that Paronychomys from the late Hemphillian of the Redington l.f. of Arizona (Jacobs 1977) is morphologically more similar to Repomys than to Peromyscus pliocenicus. It has substantial height of crown, and the third upper and lower molars are quite reduced. Repomys could be derived from Paronychomys through the increase of height of crown. The m1 of Paronychomys has a bilobed anteroconid, but the anteromedian flexid is very shallow and obliterated at the early stage of wear (F:AM 3230). The M3 of Paronychomys is E-shaped in occlusal view at a slightly worn stage (F:AM 3249), but the posterior reentrant is lost, and the anterior reentrant formed an enamel islet (AMNH 98592, the type specimen of Paronychomys lemredfieldi) as seen in R. panacaensis. The m3 of Paronychomys is S-shaped with a deep posterolabial reentrant that lasts to the base of the crown. The m3 of Repomys could be derived from Paronychomys through the reduction of this reentrant because the posterior heel of m3 in Repomys is deflected lingually due to this reentrant. However, if R. gustelyi gave rise to R. maxumi, and R. panacaensis is on a side branch as May (1981) suggested, there is a problem for the above Paronychomys-Repomys hypothesis: R. gustelyi and R. maxumi are both greatly larger than Paronychomys tuttlei (larger species of Paronychomys) or P. lemredfieldi. Also, both R. gustelyi and Paronychomys are found in late Hemphillian faunas. It is possible that the small-sized group, including R. panacaensis, R. minor, n. sp., below, and R. arizonensis are in a different lineage from that of the large-sized group, including R. gustelyi and R. maxumi. Occurrence. Rodent Ravine section: 9702, 9616, 9619; Double Butte section: 9502; Brown Bone Beds: 9620, 9621; Rodent Hill section: 9504, 9601, 9602; Limestone Corner section: 8197, in the Panaca Fm., southeast Nevada. Range. Early Blancan.
Repomys minor sp. nov.
Holotype. UALP 21369 from Locality 9504: left mandible with incisor and m1-m3. Etymology. minor, refers to small size. Material examined. UALP Locality 8197: M1s: 21161-21172, 21204; M2s: 21173-21181; M3: 21189; m1s: 21192-21200, 21137; m2s: 21206-21210; m3s: 21214-21216. Locality 9502: M1s: 21397-21399; M2: 21401. Locality 9504: left maxillary with M1-M2: 21374, 21664; M1s: 21385, 21665, 21669, 21371, 21372; M2s: 21656, 21674, 21740; M3: 21392; m2: 21383; m3s: 21696, 21698, 21699, 21396, 21760. Locality 9616: m1: 22560; m2: 22561. Locality 9620: M1: 22520; M2s: 22521, 22522; M3: 22523; m1s: 22524, 22525; m2: 22518. Locality 9621: M1s: 22548-22550; M2: 22547; m1: 22551. Locality 9601: M2: 22639; m1s: 22641, 22642; m2: 22643; m3s: 22644-22646. Locality 9602: M2: 22773; M3s: 22774, 22775; m1s: 22777, 22778; m2: 22779; m3: 22756. Locality 9702: M1s: 22824-22825, 22827-22828; M2s: 22833-22835; m1s: 22806, 22836-22838; m2s: 22840-22844; m3s: 22839, 22845.
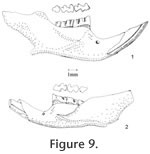
M1 is very high-crowned, as tall as long in slightly worn teeth. It has five major cusps: anterocone, protocone, paracone, hypocone, and metacone. The anterocone is asymmetrically bilobed in unworn or slightly worn specimens, with the labial conule slightly larger than the lingual. The anteromedian flexus is very shallow, and it is lost at the early stage of wear. The anterostyle on the posterior arm of the anterocone slightly constricts the paraflexus, and the mesostyle on the posterior arm of the paracone slightly constricts the metaflexus. These two small styles are obliterated at the moderately worn stage, and they never join the opposing enamel to form an islet prior to the loss of metaflexus through wear. In slightly worn specimens, the protocone is located slightly anterior to the paracone, and the hypocone is slightly anterior to the metacone. In well-worn specimens, the hypocone and metacone are nearly transversely opposed. The protocone is differentiated from the anterocone by a shallow protoflexus, and this protostria persists to just above the base of the crown. The posterior arm of the protocone may be separated from the anterior arm of the paracone in unworn specimens, but they become confluent with moderate wear. A small enterostyle is present on the posterior arm of the protocone in unworn to slightly worn specimens. A broad and deep hypoflexus separates the protocone and hypocone. The anterior arm of the hypocone is confluent with the paracone, and its posterior arm is confluent with the metacone. The paracone is separated from the anterocone by a deep, posterolingually directed paraflexus. It is separated from the metacone by a deep metaflexus. The metaflexus is directed posterolingually on unworn and moderately worn specimens and changes to a more transverse direction with wear. Both parastria and metastria continue to the base of the crown. Each M1 has three distinct roots. M2 is similar to the M1 except for its shortness and root condition. The shortness results from the loss of the anterocone. There is no anterostyle on the posterior arm of the anterocone even on young individuals, but the mesostyle between the paracone and metacone is as distinct as in M1. Each M2 has three roots (18 specimens). The anterolingual root is the strongest, and the anterolabial root is reduced. M3 is well reduced, and the occlusal outline changes dramatically at different wear stages. In unworn specimens, two labial reentrants are present so that the occlusal surface is E-shaped. The posterior reentrant is usually shallower than the anterior reentrant. The anterior cingulum is developed. The protocone is distinct, and the paracone is large. The hypocone is smaller than the protocone but is usually distinct until moderately worn. It is separated from the protocone by a shallow hypoflexus. The metacone is indistinct or absent. In slightly worn specimens, the hypocone and posterior cingulum are reduced, forming a rounded posterior heel. With wear, the posterior cingulum connects to the paracone closing the metaflexus to form a smaller and shallow posterior enamel islet. The enamel wall of this islet is very thin, as that of R. panacaensis. With wear, the paracone may connect to the well-developed anterior cingulum to form a large and shallow anterior enamel islet that may persist until moderate wear. With greater wear the anterior and posterior islets are lost. Of the four M3s, two have two roots, and two have one root. The mandible is more slender than that of R. panacaensis. The diastema between the incisor and m1 is long relative to the mandible depth (Figure 9). Its ratio is 1.28, whereas it is 0.99 in R. panacaensis. The incisor is 0.80 mm wide, and the mandible depth below m1 is 3.20 mm. The alveolar length of m1-m3 is 4.20 mm. All of these measurements are outside the respective ranges of R. panacaensis. The capsular process for the reception of incisor is less prominent than in R. panacaensis. The m1 is hypsodont, as tall as long in unworn specimens. It has five cusps: anteroconid, protoconid, metaconid, entoconid, and hypoconid. The anteroconid is single-cusped with an anterolabial cingulum directed posterolabially. The anterior cingulum is lost after moderate wear. The metaconid is distinct but smaller than in R. gustelyi and R. maxumi. It confluently connects to the anteroconid and protoconid. The m1 has a slightly different occlusal pattern from that of R. panacaensis. At the similar wear stage, the entoflexid is deeper and pointed more anteriorly in R. minor, which results in the metaconid being further from the anteroconid. The anterior arm of the protoconid connects to the anteroconid, and the posterior arm is confluent with the anterior arm of the entoconid even in unworn specimens. The anterior arm of the entoconid joins a small mesostylid in unworn and slightly worn specimens, slightly constricting the apex of the entoflexid. The hypoconid and protoconid are labially separated by a broad and moderately deep hypoflexid, with the apex directing anterolingually. The anterior arm of hypoconid confluently joins the labial side of entoconid. The protostriid is very shallow, and the hypostriid is deep but well above the base of the crown. The entostriid is slightly deeper than the posterostriid. The lingually directed posterior cingulum is very well developed and is broadly confluent with the hypoconid. The posteroflexid is slightly shallower and much narrower than the entoflexid. The m1 has two distinct roots: anterior and posterior. The m2 is similar to m1 except it is shorter. The anterior cingulum is relatively broad, with a short labial extension anterior to the protoconid. The entoflexid is directed anterolabially in all specimens. The posterior cingulum is longer in unworn and slightly worn specimens than in well-worn specimens. The enamel wall on the anterior side of the posterior cingulum is very thin in unworn and slightly worn specimens, like that of R. arizonensis. The protostriid is very shallow; the hypostriid and entostriid are very deep but terminate above the base of the crown; the posterostriid is very shallow. The m2 has two distinct roots as in m1. The m3 is reduced relative to m1 and m2. It is keyhole shaped in occlusal view in moderately worn specimens. In unworn specimens, a shallow entoflexid is present, with the apex pointing anterolingually. An anterolabial cingulum is present in slightly worn specimens and absent in moderately worn specimens. The posterior heel represents the greatly reduced hypoconid and is deflected lingually. The posterolabial reentrant (hypoflexid) is very broad and shallow, as is the posteroflexid. The entostriid is very shallow; the posterostriid and hypostriid are deeper, with the hypostriid nearly reaching the base of the crown. Of the 12 m3s, five have only one root.
Repomys minor occurs with R. panacaensis in the same localities in the Fm. except for Locality 9619 in Rodent Ravine section. The absence of R. minor from Locality 9619 is probably due to the small sample of sediments processed from this site, rather than representing the absence of this species, as R. minor was still found from sites at a higher stratigraphic level. Except for Locality 8197 in the Limestone Corner section and Locality 9502 in the Double Butte section, the other localities produce smaller abundance of R. minor than that of R. panacaensis, especially the three localities in the Rodent Hill section. Interestingly, in the Rodent Hill section, the ratio of specimens of R. minor relative to R. panacaensis decreases as the stratigraphic levels of sites increase. This pattern is also observed in the Rodent Ravine section and Brown Bone Beds (Table 8), which could suggest a gradual replacement of R. minor by R. panacaensis in the higher stratigraphic level. Localities 8197 (ratio of R. minor to R. panacaensis is 211%) and 9502 (ratio is 400%) are exceptions. They are relatively high stratigraphically, but R. minor seems more abundant than R. panacaensis at these sites. The ratio of specimens from Locality 9502 is probably biased by the small sample size (five specimens total, two species). Occurrence. Rodent Ravine section: 9702, 9616; Limestone Corner section: 8197; Double Butte section: 9502; Brown Bone Beds: 9602, 9621; Rodent Hill section: 9504, 9601, 9602, in the Fm., southeast Nevada. Range. Early Blancan.
Subfamily ARVICOLINAE Gray, 1882
Five stages of wear were used: young—crown not worn, roots not yet developed; slightly worn—crown slightly worn, roots developed but short; moderately worn—crown worn down about halfway, roots long but not closed; worn—crown worn down more than halfway, roots closed; well worn—crown almost worn out, roots closed.
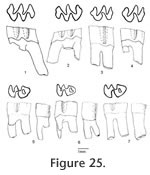
Genus MIMOMYS Forsyth Major, 1902
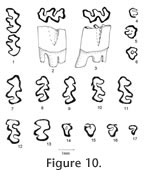
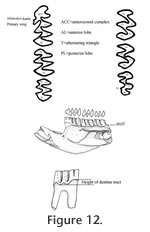
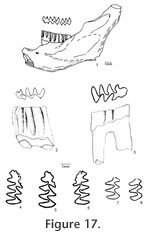
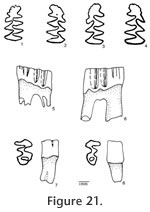
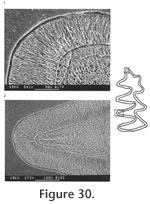
Description. UALP 22401 is an owl pellet with a partial skull, two dentaries, and several post-cranial bones. This preservation allows the direct correlation of upper and lower dentition. The upper incisor is 1.60 mm wide, without any groove or depression. The diastema between the upper incisor and M1 is 8.9 mm. The width of the two nasal bones at the widest position is 3.7 mm. The M1 has an anterior lobe and four alternating triangles (T1, T2, T3, and T4 in Figure 12,). The apices of labial reentrant angles are flexed posteriorly in moderately worn and well worn specimens. No anterior shelf is present on the M1s from Locs. 9619 and 9702, but one (out of 13) from Locality 9502 and one (out of 3) from Locality 9602 have this anterior shelf. The dentine tract is weakly developed on the lingual side of Triangle 1. Each M1 has three roots, including the anterior, lingual, and posterior roots. The M2 has an anterior lobe and three alternating triangles (T2, T3, and T4 in Figure 12). The lingual reentrant apex is slightly curved posteriorly. No reentrant pit occurs at the base of the anterior lobe. The dentine tract is weakly developed on the lingual side of anterior lobe. The majority of M2s (91%) have three roots, two anterior and one posterior. The lingual-anterior root is wide and strong. Two M2s from Locality 9619 and one from Locality 9602 have only two roots. On these specimens, the two anterior roots are fused, forming an L-shaped root.
The posterior part of the dentaries of this individual is lost, but UALP 22406 from the same locality (Locality 9702) has an almost complete dentary. It is an adult with three molars. The lower incisor passes underneath the m2 and labial relative to the m3. It ends at the labial side of the dentary between the coronoid process and condyle, forming a prominent capsular process. The mental foramen in the lower jaw is anterior to the first root of m1 on the dorsolateral surface of the dentary, slightly anterior relative to the anterior end of the masseteric crest. This position is fairly consistent in all individuals. For the measurements of lower incisor, depth of dentary, and alveolar length of m1-m3, see Table 10. The m1 has a posterior lobe, three alternating triangles, and an anteroconid complex (ACC). Labial triangles are slightly smaller than lingual triangles. The enamel is uniform in thickness. The degree of closure between the posterior lobe and the first triangle is usually slightly open; it is moderately open to open between the first and second triangles, moderately open between the second and third triangles, and slightly to moderately open between the third triangle and ACC. The apex of the second lingual reentrant is constricted and curved anteriorly. An enamel islet is usually present on unworn, slightly or moderately worn m1s. In younger specimens, the islet is larger and its shape is oval, whereas in older ones, the islet is smaller and circular. This islet disappears at the worn stage. Two m1s at the worn stage from Locality 9502 and one from Locality 9602 still preserve this islet. The "Mimomys kante" is present in all specimens except the well-worn ones. The enamel folds on the anterior-labial enamel wall only extend halfway down the crown so that the anterior ACC is quite smooth on moderately and well-worn m1s. The dentine tract on the labial side of the ACC is incipient . The m2 has a posterior lobe and four alternating triangles. Labial triangles are slightly smaller than lingual triangles. The degree of closure between alternating triangles is moderately open. The dentine tract is absent labially. The m3 has a posterior lobe and four alternating triangles, with labial triangles slightly smaller than lingual triangles. Triangle 4 is reduced and confluent with Triangle 3. The apices of lingual reentrant angles are wide. No posterior shelf is present on the m3s from Locs. 9619 and 9702. One m3 (out of five) from Locality 9502 has a posterior shelf.
Notice that M. panacaensis from these localities, especially the population from Locality 9619, has a higher percentage of M3s with two roots than the one from the type locality which has 57% of two-rooted M3s (Mou 1997). The ancestor of Mimomys, Promimomys, has three-rooted M3s; it is likely that there will be some fluctuations on the percentages of root condition of M3 in different populations of M. panacaensis. Higher percentages are not surprising due to the smaller samples from the above localities. Locality 9619 has only four M2s that preserved roots. Although all four M2s have two roots, which is the same percentage as that of M. (Ophiomys) mcknighti from the White Bluffs l.f. of Washington (Gustafson 1978), it is noticed that this population has a significant portion of M3s with enamel islets. An enamel islet was not seen in any of the six known M3s of M. (Ophiomys) mcknighti. Therefore, I refer the arvicolines from Locality 9619 to M. panacaensis. The finding of a partial skull with incisors make it possible to verify the absence of a groove on the upper incisor of M. panacaensis. This character can help us differentiate M. panacaensis from M. (Ogmodontomys) sawrockensis from the Saw Rock Canyon l.f. of Kansas, which has a groove on the upper incisor (Hibbard 1941). Other morphological differences occur in the enamel islet on m1, root condition on M3, and schmelzmuster (Mou 1997). In the dentary of M. panacaensis, the capsular process for the reception of the lower incisor is prominent, similar to that of M. (Cosomys) primus. However, several characters, including the number of roots on M3, the shape of ACC on m1, and the schmelzmuster, can readily differentiate M. panacaensis from M. (Cosomys) primus (Mou 1997, 1998). M. panacaensis is recorded in the lowest level of the Rodent Ravine section and continued to the highest level in the Rodent Hill section. Occurrence. Rodent Ravine section: 9702, 9619; Double Butte section: 9502; Rodent Hill section: 9601, 9602, 9609; Limestone Corner section: 8197, in the Fm., southeast Nevada. Range. Early Blancan.
Subfamily PROMETHEOMYINAE Kretzoi, 1955 Diagnosis. Hypsodont, rooted, prismatic cheek teeth, lacking cement in reentrant angles. The three alternating triangles on m1 have labial triangles distinctly smaller than lingual triangles. An enamel islet on ACC is only present on very young individuals and disappears in early stage of wear. The enamel folds on the anterior margin of ACC are well developed and persist into late wear. Dentine tract on the labial side of molars is very low. M3 has a large posterior islet. An anterior islet is present or absent on M3. Primitive radial enamel is present on both leading and trailing edges of alternating triangles. Differential diagnosis. The cheek teeth of Nevadomys can be distinguished from those of Promimomys by their larger size, higher dentine tract, and much less frequent retention of enamel islet on ACC of m1. Nevadomys differs from Mimomys by reduction of an islet on ACC of m1 and the much more frequent occurrence of large posterior enamel islet on M3. It is distinguished from Pliopotamys and Pliophenacomys by its smaller size, having three rather than five alternating triangles on m1, and usually having the posterior islet on M3. Genotype. Nevadomys fejfari. Etymology. Nevad, for the state of Nevada where the species ocurrs, plus mys, for mouse.
Nevadomys fejfari gen. et sp. nov.
Holotype. UALP 21358, left mandible with m1-m2. Type locality. UALP Locality 9504, in the Rodent Hill section. Etymology. fejfari, in appreciation for Dr. Oldrich Fejfar who found the type locality of this species, and for his great achievements on European arvicoline rodents.
The M2 has an anterior lobe and three alternating triangles. The lingual and second labial reentrant apices are slightly flexed posteriorly. Closure between alternating triangles as follows: slightly open between the anterior lobe and Triangle 2, slightly or moderately open between Triangle 2 and Triangle 3, and open between Triangle 3 and Triangle 4. The reentrant pit is absent on the lingual side of anterior lobe. Enamel bands have the same thickness on the occlusal surface. A dentine tract is weakly developed on the lingual side of anterior lobe. Each M2 has three roots (Table 11), two anterior and one posterior. The lingual-anterior root is wide and strong.
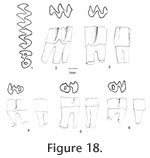
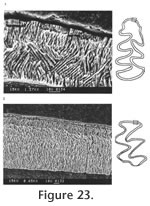
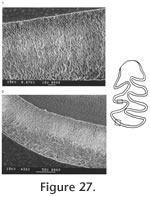
The palatal groove is as deep as that of M. (Ogmodontomys) sawrockensis, deeper than that of M. panacaensis. The mental foramen in the lower jaw is anterior to the root of m1 on the lateral surface of the dentary, about the same height as the apex of the masseteric crest. Its position is consistent in young and old individuals. The incisor passed laterally under m3, ending on the labial side of the ramus between the coronoid process and the condyle. In young individuals (UALP 21358), the capsular process is only an indistinct swelling, but in old individuals (UALP 21646), the process is prominent. The m1 has a posterior lobe, three alternating triangles, and an ACC. Labial triangles are much smaller than lingual triangles. The degree of closure between the alternating triangles is the same as that of M. panacaensis: slightly open between the posterior lobe and Triangle1, moderately open between Triangle 1 and Triangle 2, slightly or moderately open between Triangle 2 and Triangle 3, and slightly or moderately open between Triangle 3 and ACC. The apex of the second lingual reentrant angle curves anteriorly. "Mimomys kante" is absent on two-thirds of m1s (Table 12). The enamel folds on the anterior-labial surface of ACC are well developed and extend quite low in depth, close to the base of the enamel (Figure 17.4-17.6). The enamel islet is much reduced compared to that of M. panacaensis. There is no islet on any moderately or well-worn specimens. Only young individuals show this structure, and it disappears after a slight wear (Table 12). The islet is small and circular. The dentine tract on the labial side of ACC is incipient, comparable to that of M. panacaensis (Table 12). The m2 has a posterior lobe and four alternating triangles. Labial triangles are slightly smaller than lingual triangles. The degree of closure between alternating triangles is moderately open. The dentine tract is absent labially. In young individuals, enamel is thinner on the anterior surface of Triangle 4 (Figure 17.7), but the rest of enamel band is uniform in thickness. The posterior shelf is absent on m2. The m3 has a posterior lobe and four alternating triangles, with two deep lingual reentrants and two shallow and broad labial reentrants. Labial triangles are slightly smaller than lingual triangles. Triangle 4 is reduced and confluent with Triangle 3. As in the m2s, enamel is thinner on the anterior surface of Triangle 4 of unworn or slightly worn specimens. The posterior shelf is absent on m3. Four molars, including two m1s, one M1, and one M3 were chosen for examination of their schmelzmuster of enamel bands. These molars differ from M. panacaensis in that, the leading edge of ACC of m1 is occupied completely by primitive radial enamel, and no discrete lamellar enamel is seen anywhere even at the apex of alternating triangles (Figure 19.1). The leading edges of alternating triangles of all specimens have two radial enamel bands with about the same thickness, which is similar to that of M. panacaensis. The inner radial enamel (close to dentine) is more compressed than the outer radial enamel (Figure 20). However, the trailing edges of alternating triangles are also composed of two radial enamel bands, similar to the schmelzmuster of the leading edges. Primitive tangential enamel is only present on the trailing edge of the posterior lobe of m1 (Figure 19.2) and the anterior lobe of upper molars. Some primitive tangential enamel can also be observed on the trailing edges of Triangle 2 of m1s. Discussion. Nevadomys fejfari can be easily distinguished from North American arvicoline rodents by the shape of ACC on m1 and the large posterior enamel islet on M3. Enamel folds on the anterolabial side of ACC are well developed and extend down close to the base of the enamel. Enamel islet on ACC is much reduced, and the disappearance of this islet occurs at an earlier stage of wear than that of M. panacaensis, M. (Cosomys) primus, or species of M. (Ophiomys), in which the islet persists until the tooth is moderately worn. Reduction of the enamel islet is also seen in the species of M. (Ogmodontomys) (Zakrzewski 1967) and Pliophenacomys, but N. fejfari can be readily distinguished from M. (Ogmodontomys) by the presence of enamel folds on its ACC and its distinctive M3. Pliophenacomys has five alternating triangles on m1, and the enamel folds on ACC are better developed on the labial side. The posterior islet on M3 in Pliophenacomys is absent. Nevadomys fejfari has a higher dentine tract on the labial side of ACC than Promimomys or Microtoscoptes; its dentine tract is comparable to M. panacaensis and M. (Ophiomys) mcknighti. Compared to M. panacaensis, on one hand, N. fejfari has dental characters more derived in (1) reduction of enamel islet on ACC and (2) reduction of root numbers on M3; on the other hand, N. fejfari has some characters more primitive in the schmelzmuster of enamel bands and the enamel islet on M3. The thinner enamel on the anterior surface of m2 and m3, and on the posterior surface of upper molars that is characteristic of N. fejfari is not observed in M. panacaensis. However, this enamel thinning differs from the concept of enamel differentiation that we observe in advanced arvicolines, as the leading edges and trailing edges in these molars of N. fejfari have the same thickness. N. fejfari is less complex and probably more primitive than that of M. panacaensis. It has no traces of lamellar or discrete lamellar enamel anywhere in the enamel band. The leading and trailing edges of alternating triangles are composed of radial enamel. The primitive tangential enamel on the posterior lobe of m1 and the anterior lobes of M1 and M3 is not developed as well as that in M. panacaensis. This population has some characters similar to European Mimomys vandermeuleni (Fejfar et al., 1990): 1) There are deep and well-developed fine enamel folds on the anterior margin of ACC. 2) The M3 has a large posterior islet, and the anterior islet is also frequently present. 3) The M3 has two roots. 4) Size is similar. 5) Dentine tract on the labial side of ACC has similar height. However, M. vandermeuleni has a deep and large enamel islet on ACC of m1. Its descendant in Europe, M. davakosi, retains this character. Since N. fejfari has so many distinctive characters compared to M. panacaensis as well as the other known Mimomys, I consider it a new genus, and it is probably native to North America. For a phylogenetic discussion, please see the section "Remarks on Nevadomys" below Occurrence. Rodent Hill section: 9504, in the Fm., southeast Nevada. Range. Early Blancan.
Nevadomys cf. N. fejfari Material examined. UALP
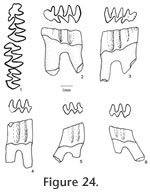
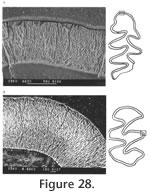
Description. The M1 has an anterior lobe, three alternating triangles, and a posterior lobe. An anterior shelf is present on 18% (3 out of 17) of the observable specimens. One M1 has an enamel interruption on the posterior surface of Triangle 4 (UALP 21894). The M2s are similar to those of N. fejfari from the type locality. Each M2 has three roots. As seen on the M3s of N. fejfari from the type locality, the majority of M3s (88%) from Locality 9620 have a large posterior enamel islet, and each tooth has only two roots. The teeth that do not have a posterior enamel islet have the Triangle 3 bending posteriorly. This bending is not seen in any M3 of M. panacaensis. The m1 has a posterior lobe, three alternating triangles, and an ACC. As in the m1s from the type locality of N. fejfari, the enamel islet is only present on young individuals (Table 12; Figure 21.1-21.4), and fine enamel folds are well developed on the anterior and anterolabial surfaces of ACC. The degree of closure between alternating triangles is similar to that of N. fejfari from the type locality. The m2s and m3s are similar to N. fejfari, respectively. The enamel on the anterior surface of Triangle 4 is thinner than the rest of the enamel band on unworn or slightly worn m2s and m3s, but in more worn specimens, the enamel thinning is absent. The posterior shelf is absent in all m2s and m3s.
Two m1s and one M1 are investigated for schmelzmuster. On one m1, a well developed, thick lamellar enamel layer is present on the leading edge of ACC (Figure 23.1), but on the other m1, a thick discrete lamellar enamel layer is observed. The leading edges of alternating triangles are composed of two radial enamel layers, similar to those of N. fejfari from the type locality, and the trailing edges have the same schmelzmuster as the leading edges with the exception that primitive tangential enamel is observed on the trailing edges of the posterior lobe of m1s and anterior lobe of the M1 (Figure 23.2).
Occurrence. Brown Bone Beds: 9620, in the Panaca Fm., southeast Nevada. Range. Early Blancan.
Nevadomys lindsayi gen. et sp. nov.
Holotype. UALP 21809: an old adult with left and right mandibles with m1-m3. Type locality. UALP Locality 9616, in the Rodent Ravine section. Etymology. lindsayi, named for Dr. Everett H. Lindsay for his outstanding contributions to the research of North American rodents. Material examined. UALP Locality 9616: M1s: 21762-21776; M2s: 21777-21789; M3s: 21790-21808; m1s: 21810-21824; m2s: 21825-21839; m3s: 21840-21848. Description. The M1 has an anterior lobe and four alternating triangles. The apices of labial reentrant angles are flexed posteriorly in moderately well worn specimens. The degree of closure between the anterior lobe and Triangle 1 is slightly open, open between Triangle 1 and Triangle 2, slightly open between Triangle 2 and Triangle 3, and open between Triangle 3 and Triangle 4. The enamel is thinner on the posterior surface of Triangle 4 on a slightly worn specimen (UALP 21771). Other enamel bands have the same thickness. No anterior shelf is seen on the M1s. Each M1 has three roots, including the anterior, lingual, and posterior. The M2 has an anterior lobe and three alternating triangles. The lingual reentrant apex is slightly curved posteriorly. The degree of closure between the anterior lobe and Triangle 1 is slightly open, slightly to moderately open between Triangle 1 and Triangle 2, and open between Triangle 2 and Triangle 3. The dentine tract is weakly developed on the lingual side of anterior lobe. No reentrant pit is seen on the M2s. Each M2 has three roots (Table 11), two anterior and one posterior. The lingual-anterior root is wide and strong.
The m1 has a posterior lobe, three alternating triangles, and an anteroconid complex. Its size is significantly larger than N. fejfari or N. downsi. Only on two juvenile m1s is there an enamel islet on ACC. The islet is small and circular, like those of N. fejfari. The degree of closure between alternating triangles is slightly open to moderately open. The apex of the second lingual reentrant is curved forward. "Mimomys kante" is absent on each m1. The fine enamel folds are well developed on the anterior margin of ACC, and they disappear at the worn stage. The dentine tract on the labial side of ACC is incipient, similar to that of N. fejfari (Table 12).
The m3 has a posterior lobe and four alternating triangles, with labial triangles distinctly smaller than lingual triangles. The degree of closure between the posterior lobe and Triangle 1 is slightly open or moderately open, open between Triangle 1 and Triangle 2, slightly or moderately open between Triangle 2 and Triangle 3, and open between Triangle 3 and Triangle 4. As in the m2s, thinner enamel is present on the anterior surface of slightly worn specimens (Figure 24.5). The posterior shelf is absent on m3.
Since one of the differences of Nevadomys lindsayi from N. fejfari is the three-rooted M3s, UALP 21794 was chosen to represent M3 schmelzmuster. It presents the same pattern as that of N. fejfari. One-third of the enamel thickness of primitive tangential enamel is present on the trailing edge of the anterior lobe. The leading and trailing edges of alternating triangles are composed of radial enamel.
The schmelzmuster of Nevadomys lindsayi is primitive in that no trace of discrete or true lamellar enamel is seen on any part of the enamel band, and primitive tangential enamel is poorly developed. This pattern is very similar to that of N. fejfari. A slight difference from N. fejfari is that N. lindsayi has true tangential enamel on the trailing edge of posterior lobe of m1s. Locality 9504, where Nevadomys fejfari came from, is stratigraphically higher than Locality 9616. N. fejfari could be derived from N. lindsayi by the fusion of two anterior roots on M3 and decreasing in overall size. Occurrence. Rodent Ravine section: 9616, in the Panaca Fm., southeast Nevada. Range. Early Blancan.
Nevadomys downsi gen. et sp. nov.
Holotype. UALP 22054, a left m1. Type locality. UALP Locality 9621, in the Brown Bone Beds section. Etymology. downsi, in appreciation for the late William R. Downs who found several localities in the Fm. Material examined. UALP Locality 9621: right maxillary with M1-M2: 22091; right M2-M3: 22028; M1s: 21996-22010; M2s: 22011-22027; M3s: 22029-22046; right mandible with m1-m2: 22047; right mandible with m1: 22048; left m1-m2: 22049; m1s: 22050-22064; 22065-22076; m3s: 22077-22090.
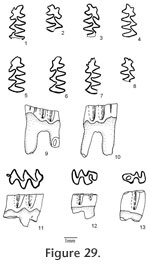
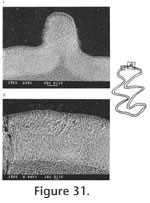
The M2 has an anterior lobe and three alternating triangles. Lingual triangles are slightly smaller than labial triangles. The lingual reentrant apex is slightly curved posteriorly. The degree of closure between the anterior lobe and Triangle 2 is slightly open; it is slightly open between Triangle 2 and Triangle 3, and moderately open between Triangle 3 and Triangle 4. The dentine tract is incipiently developed on the lingual side of anterior lobe. No enamel prism is observed on the anterior surface of M2. Each M2 has three roots (Table 11), two anterior and one posterior. The lingual-anterior root is wide and strong. The M3 has an anterior lobe, three alternating triangles, and a posterior lobe. Labial triangles and reentrants are much smaller than lingual ones. Seventy-nine percent of M3s have the posterior islet. Three specimens (UALP 22033, 22037, and 22038) have the islet incompletely closed by having the posterior lingual reentrant slightly open. The islet will be formed with further wear. Four specimens (UALP 22031, 22032, 22034, and 22035) lack the posterior islet and have the posterior lingual reentrant open until late wear. Within these four specimens, three have a posteriorly bent Triangle 3 (Figure 29.12), as some M3s of Nevadomys cf. N. fejfari from Locality 9620. One (UALP 22032) out of 19 specimens has a small anterior islet. As in N. fejfari, the majority of M3s (88%) have only two roots. The m1 from this site has a posterior lobe, three alternating triangles, and the ACC. Labial triangles are distinctly smaller than lingual triangles. The enamel folds are well developed on the anterior margin of ACC, but they disappear at the moderately worn stage. After the crown is slightly worn, a single narrow but deep fold is preserved on the anterior margin of ACC. A labial fold persists on the ACC until moderate wear. The ACC do not have "Mimomys kante." An enamel islet is present only on very young individuals. The degree of closure between the posterior lobe and Triangle 1 is slightly open; it is open between Triangle 1 and Triangle 2, moderately open between Triangle 2 and Triangle 3, and slightly to moderately open between Triangle 3 and the ACC. The dentine tract is gently rounded, slightly higher than N. fejfari from Locality 9504 and 9620 (Table 12).
The m3 has a posterior lobe and four alternating triangles, with labial triangles much smaller than lingual triangles. The degree of closure between alternating triangles is slightly open between the posterior lobe and Triangle 1, moderately open between Triangle 1 and Triangle 2, slightly open between Triangle 2 and Triangle 3, and open between Triangle 3 and Triangle 4. As in the m2s, thinner enamel is present on the anterior surface of unworn or slightly worn specimens. No posterior shelf is present on any m3. Two m1s and one M1 were examined under SEM for their schmelzmuster. The leading edge on ACC of two m1s is composed of radial enamel. A few prisms close to the dentine at the apices of the lobes on ACC may extend toward the occlusal surface, forming a trace of discrete lamellar enamel (Figure 30.1). The leading edges and trailing edges have the same pattern, and both are composed of two radial enamel bands. The inner layer is more compressed than the outer layer (Figure 30). Primitive tangential enamel is observed on the trailing edge of the posterior lobe of m1. No discrete lamellar enamel is seen on the apices of the alternating triangles. A moderately worn M1 (UALP 21999) with a distinct enamel "prism" on the anterior surface was chosen for examination. Traces of primitive tangential enamel are seen on the trailing edge of the anterior lobe, and the enamel "prism" on the anterior surface is composed totally of radial enamel (Figure 31). Discussion. This population has the following characters similar to Nevadomys fejfari: 1) size; 2) enamel folds on the anterior margin of m1; 3) reduction of enamel islet on m1; 4) two-rooted M3; 5) majority of M3s with the posterior islet; and 6) primitive radial enamel. However, it also has several characters that are different from N. fejfari: 1) the four-lobed ACC that is caused by a deep prism fold on the anterior and lingual surfaces; 2) absence of "Mimomys kante" on majority of m1s; 3) slightly higher dentine tract on the labial side of ACC; 4) a strong but narrow enamel prism may be present on the anterior surface of M1; and 5) incomplete closure of the posterolingual reentrant in M3 of some specimens. Locality 9621 is only 2 m higher than Locality 9620 in the same section (Brown Bone Bed section). The latter produced a population identified as close to Nevadomys fejfari. These two species probably evolved from their ancestor somewhere else in North America and immigrated to Panaca at about the same time. They represent a similar evolutionary stage of development in the Nevadomys lineage. The four-lobed ACC on m1 is also seen in some European arvicolines: Germanomys, Stachomys, and Ungaromys (Fejfar et al. 1998). Stachomys and Ungaromys are distinctive from Nevadomys in their m2, m3, and M3 (Fejfar et al. 1998, plate 7). Germanomys (Fejfar and Repenning 1998) is more similar to Nevadomys in the following characters: 1) well-developed fine enamel folds on the anterior of ACC of m1; with further wear, the ACC is four-lobed; 2) reduction of enamel islet on ACC; 3) comparable low dentine tract on the labial side of ACC; and 4) the Triangle 3 on M3 bent posteriorly, as in some M3s of Nevadomys fejfari and N. downsi (percentages or root condition is not published). However, the closure degree between Triangle 3 and ACC is more open than that of N. downsi, and there is a distinct reentrant on Triangle 4 of m2 that is not seen in the m2 of N. downsi. A shallow depression, instead of a distinct enamel prism, is present on the anterior surface of M1. Based on these differences, I tentatively consider that the population from Locality 9621 is a species of Nevadomys rather than Germanomys. Occurrence. Brown Bone Beds: 9621; in the Fm., southeast Nevada. Range. Early Blancan.
Remarks on Nevadomys. To summarize the differences of Nevadomys and Mimomys panacaensis, the genotype N. fejfari is chosen to compare with M. panacaensis as the following:
Nevadomys fejfari
Mimomys panacaensis
Enamel islet on m1
not reduced (primitive)
"Mimomys kante"
present
Enamel folds on ACC
lost at early worn stage
Posterior enamel isleton M3
few(<10%) (derived)
Roots on M3
half with 3 roots (primitive)
Schmelzmuster
discrete lamellar or lamellar on CC N. fejfari have some more advanced as well as more primitive characters relative to M. panacaensis. Therefore, it is not likely that the two genera share the most recent ancestor. M. panacaensis is probably an immigrant from Europe, whereas Nevadomys is most likely endemic to North America. The earliest known species that have prismatic, high-crowned cheek teeth in North America are Goniodontomys and Microtoscoptes from the Hemphillian NALMA. Microtoscoptes was first reported from Ertemte, Inner Mongolia of China based on one m1 and M1 (this tooth was actually an M2, Fahlbusch 1987). The genotype is M. praetermissus. Its diagnosis was summarized by Gromov and Polyakov (1977) as the following: lingual triangle of M2 and M3 about half the size of the labial one; only one lingual reentrant on M2, and correspondingly, only two lingual triangles on the tooth; M3 shorter than M2; posterior lobe of M3 simple. Goniodontomys was erected by Wilson (1937) based on two lower jaws from Oregon. Wilson (1937) named the genotype species Goniodontomys disjunctus, with its diagnosis that the triangles are opposite rather than alternating. Hibbard (1959) studied similar specimens from Wyoming and Idaho, and he changed Goniodontomys disjunctus into Microtoscoptes disjunctus, considering that Goniodontomys is the junior synonymy of Microtoscoptes. This opinion was followed by Repenning (1968), Hibbard (1972), and Shotwell (1970), but was not followed by Gromov and Polyakov (1977) and Fahlbusch (1987). Comparing the Eurasian and North American specimens, Gromov and Polyakov (1977) gave a diagnosis of Goniodontomys as the following: lingual triangle of M2 and M3 the same size as the labial one, facing oppositely; two lingual reentrants on M2, and correspondingly, three lingual triangles on the tooth; M3 longer than M2; a deep reentrant on the posterior lobe of M3. Martin (1975) studied some Hemphillian arvicoline rodents from the upper portion of the Ogallala Group in Nebraska. He identified some specimens as Microtoscoptes disjunctus, but he recognized Microtoscoptes disjunctus of Shotwell (1970) and some other specimens from Nebraska as a new genus, Paramicrotoscoptes, with the new species designated as Paramicrotoscoptes hibbardi (genotype of Paramicrotoscoptes). This species is also characterized by its opposite triangles; its M2 has only one broad lingual reentrant, which is similar to that of Microtoscoptes praetermissus but different from that of Goniodontomys disjunctus. However, its M3 differs from that of M. praetermissus in its larger size and the presence of a posterior enamel islet. In his review, Repenning (1987) made the following changes informally: 1) Microtoscoptes disjunctus was changed back into Goniodontomys disjunctus, following Gromov and Polyakov (1977); 2) Paramicrotoscoptes was considered the junior synonymy of Microtoscoptes; the species Paramicrotoscoptes hibbardi was changed to Microtoscoptes hibbardi, which was the only species of Microtoscoptes in North America. This classification was later followed by Korth (1994) in his review of North American Tertiary rodents. However, almost at the same time, Fahlbusch (1987) studied a much larger sample of Microtoscoptes praetermissus (about 150 specimens) from Ertemte, the topotypic locality and described the morphology of the whole dentitions of this species. He concluded that Microtoscoptes and Goniodontomys are two distinct genera, with the major differences on the morphology of M2, M3, and m3. Paramicrotoscoptes is different from Microtoscoptes, but closer to Microtoscoptes than to Goniodontomys. Along with this paper (Fahlbusch 1987), Koenigswald (1987) described the enamel microstructure of molars of the three genera. He found that Microtoscoptes and Goniodontomys have similar schmelzmuster, whereas Paramicrotoscoptes has a different schmelzmuster. Based on the above reviews and examination of Martin's (1975) illustration, I agree with Gromov and Polyakov (1977) and Fahlbusch (1987) that Microtoscoptes, Goniodontomys, and Paramicrotoscoptes are three distinct genera. So far, Microtoscoptes has not been found in North America, whereas Goniodontomys and Paramicrotoscoptes have not been found in Europe. Repenning (1987) derived Pliophenacomys from "Propliophenacomys," and further suggested that Paramicrotoscoptes gave rise to "Propliophenacomys" (Repenning et al. 1990). However, the intermediate genus in this lineage, "Propliophenacomys," is very poorly documented as pointed out by Korth (1994). Compared to the taxa Goniodontomys disjunctus, Paramicrotoscoptes hibbardi, and "Propliophenacomys" parkeri, Nevadomys seems more closely related to Paramicrotoscoptes hibbardi. In the m1s of P. hibbardi, the ACC is four-lobed with a deep anterior enamel fold, which is similar to that of Nevadomys downsi. The enamel islets only exist on unworn or slightly worn m1s in P. hibbardi, and Nevadomys is derived by further reduction of this islet. A large posterior enamel islet is present on the M3 of P. hibbardi; Nevadomys still retains this character, but the M3 is more derived by its reduction (as in N. lindsayi) or even fusion (as in N. fejfari and N. downsi) of the anterolingual roots. Following the phylogeny of Pliophenacomys proposed by Repenning (1987), Korth (1994) placed this genus in the same subfamily as Paramicrotoscoptes and Goniodontomys, which is Prometheomyinae. For the reasons discussed above, Nevadomys was likely derived from Paramicrotoscoptes, and is therefore placed in the subfamily Prometheomyinae. The Paramicrotoscoptes-Nevadomys relationship is not only supported by the dental morphology discussed above but also by the enamel microstructure of molars. According to von Koenigswald (1980, 1987), the leading and trailing edges of opposite triangles of Paramicrotoscoptes hibbardi has the same schmelzmuster, which is composed of two radial enamel layers, each occupying half of the width of enamel band. Microtoscoptes and Goniodontomys, which also have opposite triangles on their molars, display a different schmelzmuster from that of Paramicrotoscoptes. These two genera have the schmelzmuster composed of two radial enamel layers and a thin tangential enamel layer in the middle. Schmelzmuster of Nevadomys does not have the thin tangential enamel in the middle. Both of its leading and trailing edges of the enamel bands are composed of two radial enamel layers. It is comparable to that of Paramicrotoscoptes, but quite different from that of Microtoscoptes or Goniodontomys. |
|||||||||||||||